海尔格科尔布
1.锥信号电路。
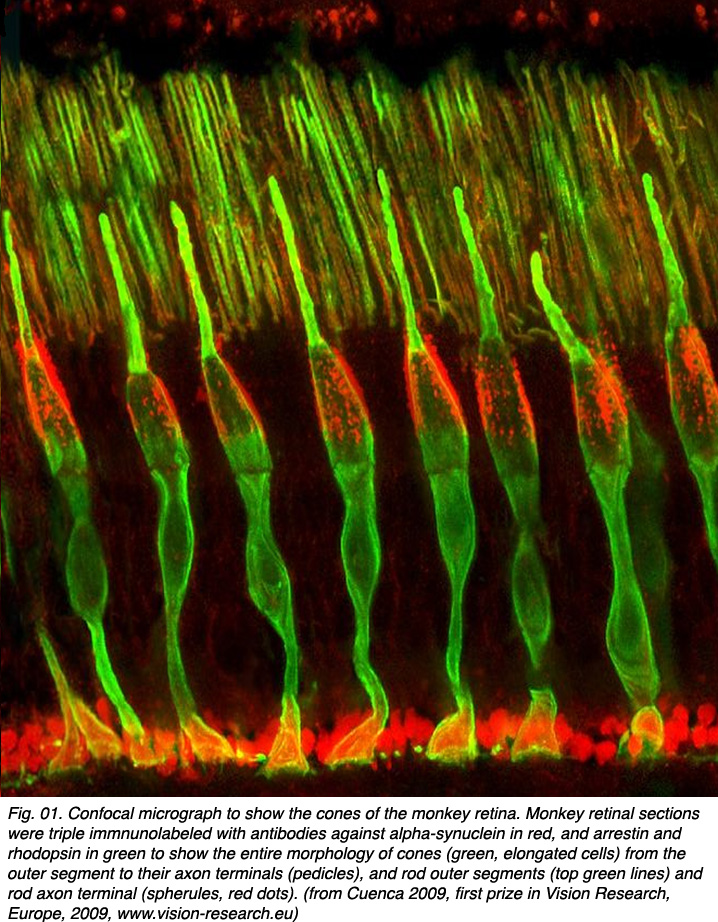
视锥光感受器是视网膜中对亮光和不同波长光的感受器。它们在光(强光)条件下是敏感的,根据视觉色素或视蛋白在其外节区域的结构分为几种类型。在双色动物中,两种视锥上有两种视色素(大多数哺乳动物)。视锥细胞对蓝光敏感,对红绿光敏感。在三色动物物种(一些灵长类动物和人类)中,根据它们的视觉色素有三种类型的锥体(见光感受器章节)。这些是长波长(红色)、中波长(绿色)和短波长(蓝色)敏感锥体。图01显示了用α -突触核蛋白和抑制素抗体染色的锥状光感受器,因此它们在猴子视网膜切片上清晰可见(由西班牙阿利坎特大学Nicolas Cuenca教授提供)。
视锥信号通过视网膜传递到神经节细胞的电路与视杆信号通路截然不同。第一个区别是在外丛状层。锥体突触在各种锥体双极类型上,而不是在单一类型上,如杆系统图02。因此,在外部丛状层已经为锥系统安装了可选择的路径。正如我们已经提到的(OPL部分),锥体双极子根据其树突场的大小(小的、弥漫的和大场弥漫的,图02)以及它们与锥体蒂接触的不同类型的突触来区分,例如,凹陷的带状突触(imb)、平坦的或基底连接(fmb)或非带状相关的基底连接(图1)。
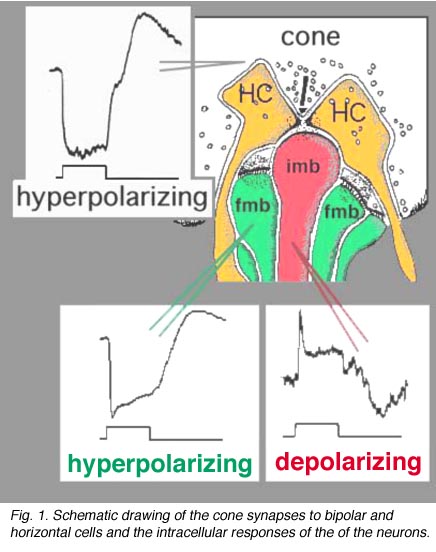
脊椎动物的光感受器在黑暗中处于去偏振状态,被光超偏振(Trifonov, 1968)。因此,人们认为神经递质谷氨酸在黑暗中持续释放,并被光抑制。我们还知道,双极细胞对光的反应要么是去极化,要么是对光刺激的超极化(Werblin和Dowling, 1969;金子,1970)。正如我们在杆状途径一章中所看到的,杆状双极细胞对光有去偏光反应(Dacheux和Raviola, 1986),但锥体双极细胞对光可以超偏光或去偏光(Nelson和Kolb, 1983)。现在,我们知道进行中心带状接触的锥体双极类型(图1,imb)将是ON-center(中心去极化)类型(ON BC),而进行宽裂基底连接的锥体双极细胞(图1,fmb)将像光感受器本身一样对光做出反应,即将是OFF-center(中心超极化)类型(Nelson和Kolb, 1983),这是Raviola和Gilula(1975)最初提出的。
不同类型的谷氨酸受体出现在去极化(on -中心)和超极化(off -中心)的双极细胞上(图2)(Miller和Slaughter, 1986)。中枢突触的ON或OFF是由双极细胞树突与锥接触处的谷氨酸突触后受体的性质决定的。off -双极受体似乎与ampa -蓝氨酸受体类型相关,因此是一种常见的兴奋性、电离性谷氨酸受体(图2,iGluR),一些是特殊的蓝氨酸受体(图2,蓝氨酸)。相反,on型双极细胞有代谢受体(mGluR),选择性结合谷氨酸激动剂APB(或ap4,2 -氨基-4-膦酸丁酸)。APB的应用选择性地提高了它们的膜电位,抑制了on中心双极细胞的光响应(Slaughter and Miller, 1981;纳威和哥本哈根,1987)。on -双极细胞上的受体现在被认为是mGluR6(图2,红色符号)(Numura et al. 1994;Vardi et al. 1997)。受体激活的g蛋白最初被认为是模拟光感受器中发生的循环- gmp级联,是on -中心双极细胞转导的潜在机制(Nawy和Jahr, 1990;谢尔斯和福尔克,1990)。 Most recently, good evidence has been provided for a subunit of the transducin molecule, GalphaO, to be the second messenger in the ON-center bipolar cell activation pathway (Nawy, 1999; Dhingra et al. 2000). See more detailed descriptions in the chapter on bipolar cells pathways by Connaughton and Nelson, webvision.
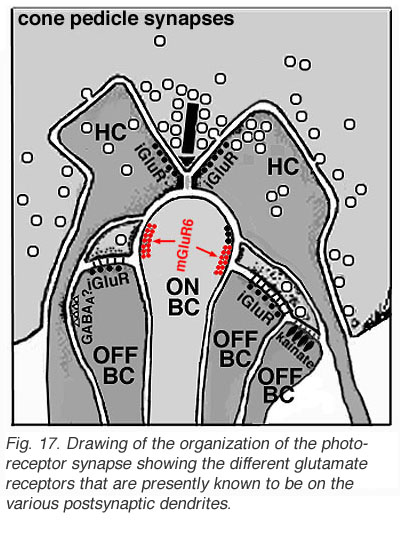
图2所示。目前对锥体传递到两极和水平细胞树突的受体分子类型的认识
几年前,通过电子显微镜和三维重建猫视网膜内丛状层的锥体双极廓形显示,这些双极轴突将大部分的带状输出突触连接到神经节细胞树突(图3)(Kolb, 1979)。
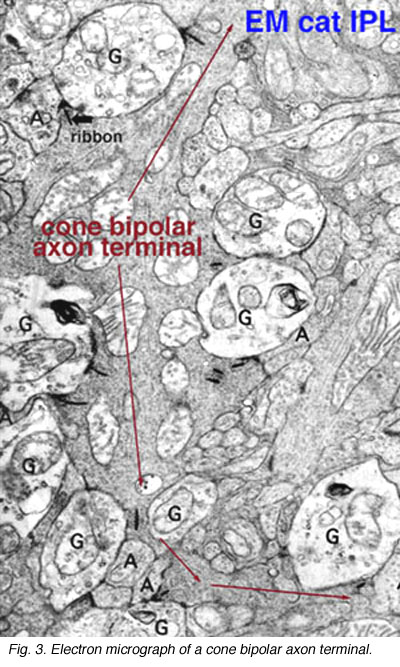 图3。内丛状层锥体双极轴突末端的电子显微图 |
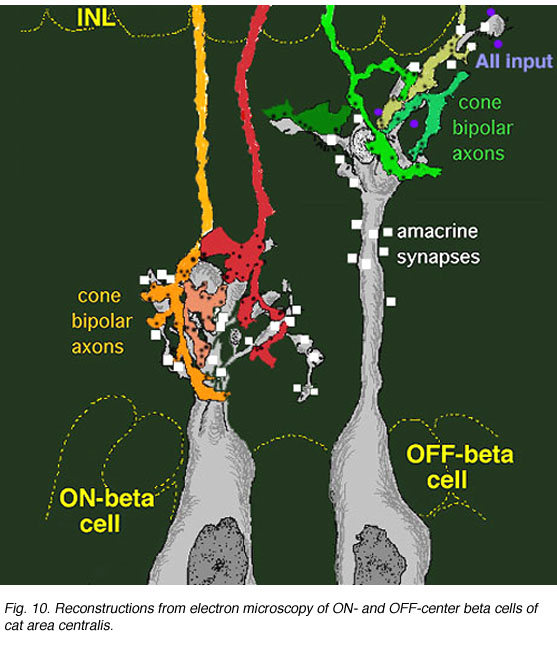 图4。锥体双极终端的三维重建及其对β神经节细胞的输入 |
两端的细胞轴突终止于亚层一个内丛状层(更接近无分泌细胞体)的神经元只与神经节细胞的树突形成带状突触,而神经节细胞的树突在这层薄板下层(图4)。事实上,这样的双极性细胞轴突甚至无法深入到足以接触到薄板下层的神经节细胞bIPL。神经节细胞在椎板下分支一个从Nelson和同事的发现(Nelson et al., 1978)中得知,对闪光产生偏离中心的反应。
相反,锥体双极细胞,轴突位于椎板下b内丛状层(靠近神经节细胞体)中,仅在节细胞的树突上形成带状突触b(图4)。Nelson等(1978)再次表明,这种神经节细胞对闪光的反应是ON-center的。因此锥体双极反应类型(ON或OFF中心)特别驱动它的ON或OFF中心神经节细胞分离到两个亚节(b而且一个分别(图4)。
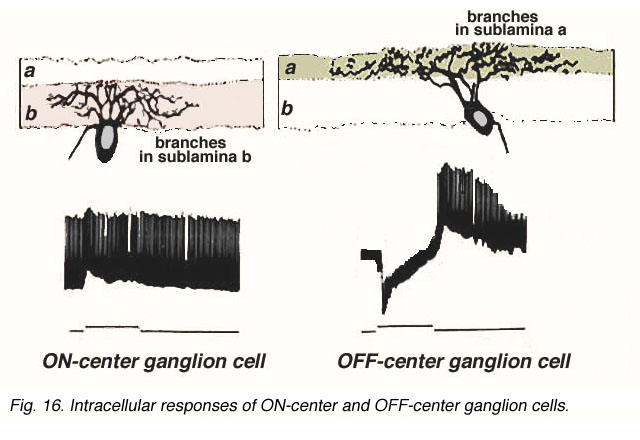
图5(下图)显示了猫视网膜内记录的和HRP染色的神经节细胞的分支水平和ON和OFF峰值反应。动画进一步说明了形态学的差异和相应的光响应。
在人类视网膜中,常见的锥体双极细胞和猫一样,不仅根据其锥体蒂突触的性质,而且根据其轴突终止于IPL的哪个亚层进行分类。因此,一些锥体双极型将轴突发送到椎板下一个(fb类型)和其他到亚鳍b(ib类型)的IPL。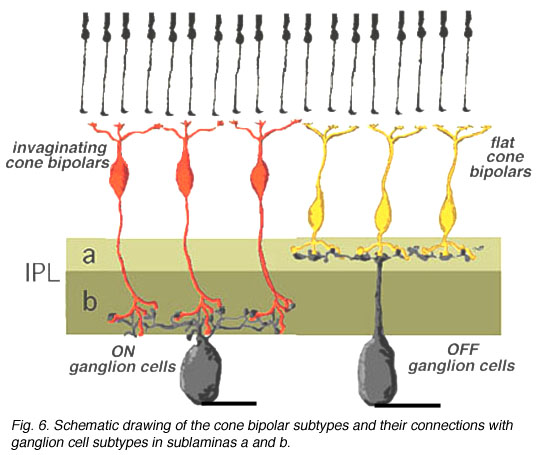 我们期望像猫一样,人的锥体双极细胞在椎板下有轴突一个将连接到OFF-center(中心超极化)神经节细胞和双极细胞与轴突在椎板下b会连接到ON-center(中心去极化)神经节细胞(图6)。
我们期望像猫一样,人的锥体双极细胞在椎板下有轴突一个将连接到OFF-center(中心超极化)神经节细胞和双极细胞与轴突在椎板下b会连接到ON-center(中心去极化)神经节细胞(图6)。
图6所示。两种锥体双极亚型(内陷型和扁平型)及其与ON和OFF中心神经节细胞的连通性概要示意图。
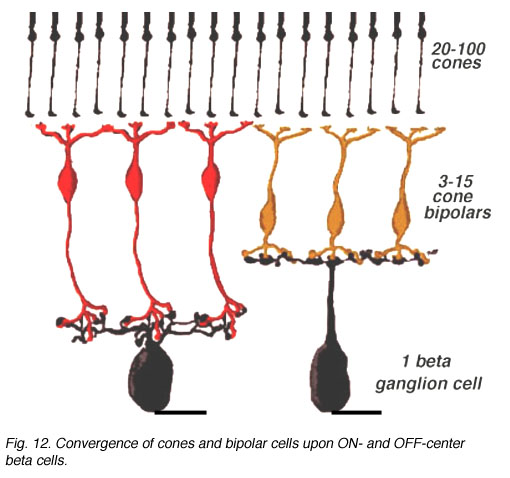
因此,与哺乳动物视网膜上的杆状细胞通路相比,锥体双极细胞与神经节细胞树突直接形成突触,而不需要杆状细胞通路中出现的中间无分泌细胞通路(见前一章)。因此,锥路径比杆路径更直接,也更窄场和收敛。锥体双极细胞聚集在锥体双极细胞上的数量比杆对杆双极细胞要少,只有相对少量的锥体双极细胞聚集在神经节细胞上(图7)。在低聚集率的最终是人类和灵长类动物视网膜中的侏儒系统,我们将在另一章单独讨论。
2.锥通路介导连续的对比(ON和OFF通路)。
哺乳动物和人类视网膜中的视锥通路是两条平行的信息流,从视锥光感受器直接通过直管道,即视锥双极细胞,到达神经节细胞。杆状系统只有一个平行通道,而锥状系统却有两个平行通道,这是什么原因?答案是,这种结构允许一个通道向神经节细胞提供比背景刺激更亮的信息(ON-center通道)和比背景刺激更暗的信息(OFF-center通道)(图8),Kuffler在1953年首次从猫视网膜神经节细胞的记录中证明了这一点。

正如我们在上面看到的双极细胞中这两个重要的ON-center通道和OFF-center通道起源的解剖学底物是锥双极细胞与锥蒂的突触接触类型以及它们与伙伴神经节细胞的特殊不同的层状接触。
通过猫的视网膜传递亮度和黑暗信息的完整电路如下图所示(图9和电影)。

3.锥体通路电路介导同时对比(中心-周围感受野)。
关于图像整体亮度或暗度的信息对于视觉感觉来说是最重要的,但是将这两个信息同时放在一起对比可以极大地提高图像的分辨率(图10)。

同时对比是通过横向抑制来实现的,其中一个暗边界抑制了一个亮区域,反之亦然。在视网膜上,Hartline(1940)从青蛙视神经记录中首次发现视网膜神经节细胞感受野是同心圆的,对感受野周围的中心有相反的响应。
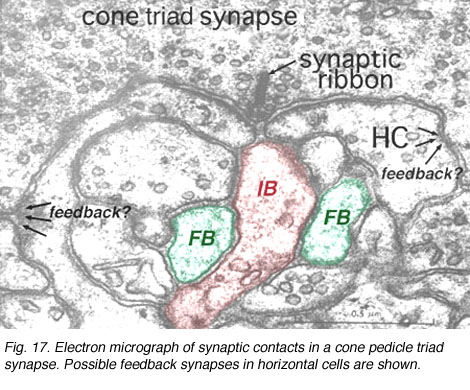
人们认为,在光下眼窝的水平细胞通过一种侧抑制机制,在感受器本身和接触光感受器的双极细胞的感受野中心周围形成一个环绕(Werblin和Dowling, 1969;贝勒等人,1971年;金子,1970)。相关的布线似乎开始于我们在带突触的锥三联体中看到的局部小电路(图11)(见外丛状层一章)。
图11所示。锥体三联征的电子显微镜图显示了水平细胞树突(HC),它们位于锥体的突触后(并可能形成对锥体的反馈突触)和两个主要的突触后双极性细胞树突(IB和FB)。
因此,水平细胞对带状三联突触的锥光感受器的负反馈突触允许水平细胞网络的更大的感受野(水平细胞通过相邻细胞之间的电突触在合胞体中跨视网膜耦合),为狭窄的中央锥反应提供包围(Werblin和Dowling, 1969;纳卡,1976)。然后,这种同心组织被传输到与锥体接触的双极细胞(Werblin和Dowling, 1969;金子,1970;Toyoda, 1972),从那里到锥体双极细胞接触的神经节细胞。
点击此处观看受刺激锥体侧抑制机制的电影(Quicktime电影)
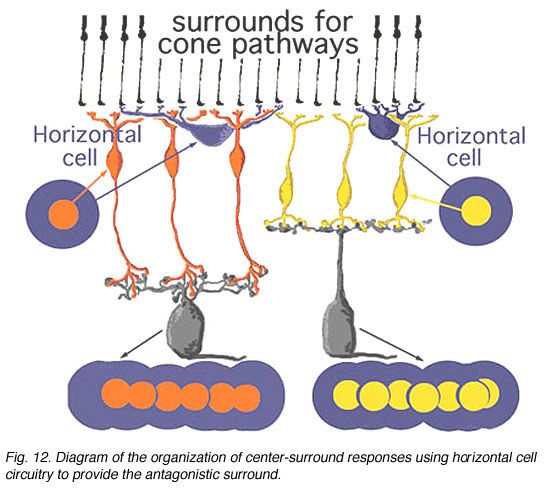
下图12总结了哺乳动物视网膜锥体双极系统中水平细胞到锥体双极细胞回路的中心环绕组织结构。中心通道是由锥体到双极到神经节细胞的通通道形成的,而水平细胞信息的注入则为中心提供了一个拮抗包围:OFF-surround用于on -中心通道(水平细胞和橙色双极,左手通路),ON-surround用于off -中心通道(水平细胞和黄色双极,右手通路)(Werblin和Dowling, 1979;Werblin, 1991)。
点击此处观看圆锥细胞、双极细胞和神经节细胞周围形成的动画(Quicktime电影)
在哺乳动物视网膜中,环绕反应在双极细胞感受野中的作用不像在冷血脊椎动物双极细胞中那样强烈(Nelson and Kolb, 1983)。下面的图13和14比较了海龟视网膜(Ammermüller and Kolb, 1995)和猫视网膜(Nelson and Kolb, 1983)的双极反应。
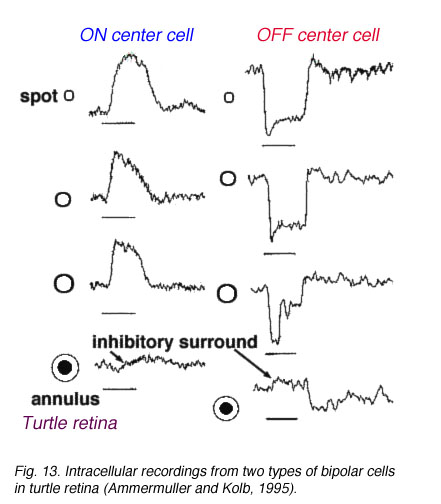 图13所示。龟视网膜双极细胞记录 |
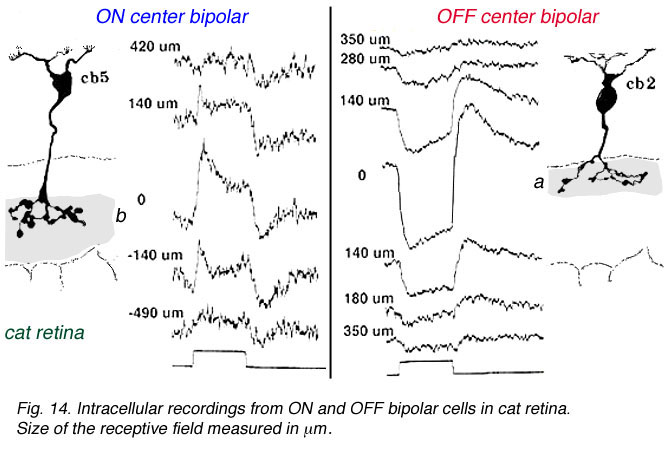 图14所示。猫视网膜的双极细胞记录 |
我们从猫和猴子神经节细胞的细胞内和细胞外记录中知道(Enroth-Cugell和Robson, 1966;Kuffler, 1953;Levick和Thibos, 1983年;Gouras, 1968;Shapley和Perry, 1986)认为最常见的哺乳动物神经节细胞具有强大的中心环绕组织。因此,对神经节细胞的双极驱动中枢反应的额外包围拮抗可能是由内部丛状层中的某些尚未完全理解的无分泌细胞网络构建的(图15)(Lebedev and Marshak, 2007)。
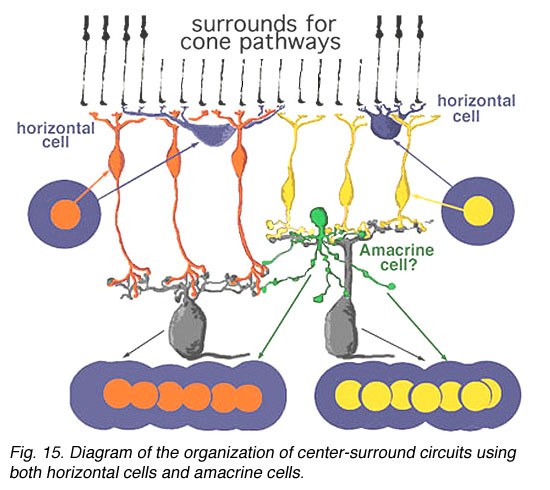
图15所示。使用水平细胞和无分泌细胞的中心环绕电路的组织示意图
4.参考文献
Ammermüller J .柯波。乌龟内视网膜的组织结构。上中心和偏中心通路。中华神经科学杂志1995;358:1-34。[PubMed]
Baylor DA, Fuortes MGF, O 'Bryan PM。海龟视网膜上视锥细胞的感受区。j .杂志。1971;214:265 - 294。[PubMed]
猫视网膜锥体双极神经元的细胞类型。生物学杂志1990;330:305-322。[PubMed]
Dhingra A, Lyubarsky A, Jiang M, Pugh EN, Birnbaumer L, Sterling P, Vardi N. ON双极神经元的光响应需要Gao。J Neurosci.2000; 20:9053 - 9058。[PubMed]
罗布森JG。猫视网膜神经节细胞的对比敏感度。杂志。1966;187:517 - 552。[PubMed]
猴神经节细胞锥体机制的鉴定。杂志。1968;199:533 - 547。[PubMed]
Hartline港元。视神经纤维的感受区。美国医学杂志。1940;130:690-699。
金鱼视网膜水平细胞、双极细胞和无分泌细胞的生理和形态学鉴定。杂志。1970;207:623 - 633。[PubMed]
猫视网膜内丛状层:电子显微镜观察。J Neurocytol。1979;8:295 - 329。[PubMed]
Kuffler西南。哺乳动物视网膜放电模式与功能组织。J Neurophysiol。1953;16:37 - 68。[PubMed]
Levick WR, Thibos LN。猫神经节细胞的感受野:分类与构建。1983; 2:267-320。
视网膜的兴奋性氨基酸受体:多样性、亚型和传导机制。趋势> 1986;9:211 - 213。
纳卡K-I。鲶鱼视网膜中的神经回路。投资。Ophthal。1976;15:926 - 935。
视网膜去极化双极细胞上的多种谷氨酸受体。大自然。1987;325:56-58。[PubMed]
Nawy S, Jahr CE。谷氨酸对视网膜双极细胞cGMP激活电导的抑制。大自然。1990;346:269 - 271。[PubMed]
在视网膜双极细胞中,代谢受体mGluR6可能通过Go发出信号,而不是磷酸二酯酶。J > 1999; 19:2938 - 2944。PubMed]
Nelson R, Famiglietti EV, Kolb H.细胞内染色显示猫视网膜中中心和非中心神经节细胞有不同程度的分层。J Neurophysiol。1978;41:427 - 483。
猫视网膜双极细胞和神经节细胞的突触模式和反应特性。视觉研究》1983;23:1183 - 1195。[PubMed]
大鼠棒状双极细胞中代谢性谷氨酸受体突触后定位的发育调控。细胞。1994;77:361 - 369。[PubMed]
沙普利R,佩里VH。猫、猴视网膜神经节细胞及其视觉功能作用。趋势> 1986;9:229 - 235。
杆状双极细胞的谷氨酸受体通过g蛋白与环GMP级联连接。生物学报1990;242:91-94。[PubMed]
斯勒特MM,米勒RF。2-氨基-4-膦丁酸:视网膜研究的新药理学工具。科学。1981;211:182 - 184。[PubMed]
丰田章男J-I。鲤鱼视网膜中双极细胞反应的膜阻力变化。视觉研究》1972;12:283 - 294。
Trifonov丫。利用视网膜电刺激光感受器和水平细胞之间突触传递的研究。Biofizika.1968; 10:673 - 680。
Vardi N, Morigawa K.大鼠ON锥双极细胞表达代谢受体mGluR6。Vis. >。1997; 14:789 - 794。[PubMed]
Werblin FS Dowling JE。泥鳅视网膜的组织,黄斑泥鳅。2细胞内记录。J Neurophysiol。1969;32:339 - 355。PubMed]
Werblin FS。虎蝾螈视网膜的突触连接,感受野和活动模式。眼科科学。1991;32:459-483 .[PubMed]
最后更新:2011年7月。