海尔格科尔布
1.一般特征。
在视网膜的每一层都有相互或反馈回路,因此某些神经元可以在同一层内横向相互作用,从一层到另一层垂直相互作用,甚至从大脑到视网膜。层内反馈环路通常是由使用抑制性神经递质的神经元提供的,如GABA,通过增加神经元的接受野的侧抑制或拮抗作用,具有锐化图像的功能,而层间或来自大脑的反馈环路在功能上不太清楚。后一种循环倾向于使用神经调质作为它们的递质,因此对神经元群或神经元链的兴奋性状态(例如适应性)有更广泛的影响。
2.视网膜外部的反馈。
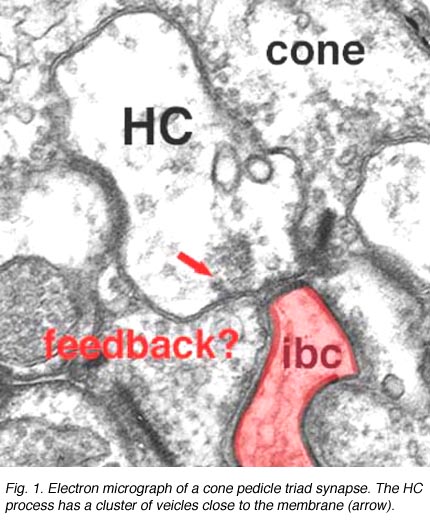 图1所示。锥体椎弓根的电子显微镜图 |
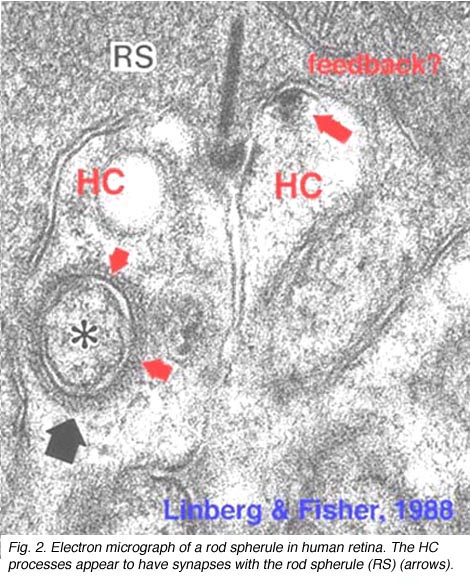 图2所示。棒状球体的电子显微照片 |
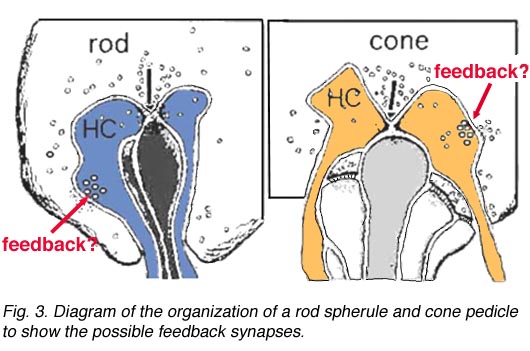
如前几章所述,在海龟和鱼的视网膜中已经很好地证明,视锥细胞从水平细胞接收到拮抗的相互反馈信息(Baylor等人,1971年),这为单个视锥细胞提供了一个受限的同心圆感受场。在哺乳动物视网膜的视锥细胞中,无论是电生理学还是形态学上都很难证明反馈突触的存在。然而,在人类视网膜中,HI水平细胞的杆状轴突末端在杆状球上(图2和图3)和杆状双极细胞树突上(图2和Fisher, 1988),可以看到在膜上单个密集投影处形成小的点状化学突触,由小簇泡囊组成(图2和图3)。在人类视网膜锥体蒂三位一体的水平细胞树突中也偶尔出现类似的小泡簇(图1和图3)。
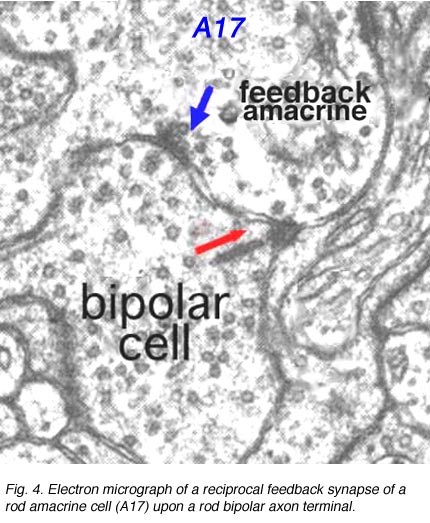
在内丛状层中,许多无分泌细胞可能向双极轴突末端提供反馈信息。其中一个我们已经了解的是来自宽视场杆状无侧突起A17在杆状双极轴突末端上的互易突触(图4)。有趣的是,杆状双极和A17无侧突起之间的IPL输入/输出突触与光感受器和OPL上水平细胞的输入/输出平行,这表明至少在哺乳动物视网膜上,杆状系统在整个视网膜上处于重复反馈循环的必要性。
图4。杆状无分泌细胞(A17)在杆状双极轴突末端上的互反突触的电子显微图
A17在哪里是a杆系统双极细胞反馈无螺孔细胞,A4, A13和宽视野无螺孔,如A19锥系统反馈无分泌细胞。像杆状系统的A17细胞一样,这些无分泌细胞可能也是GABAergic。下图所示的无分泌细胞反馈环路(图5)通常发生在人类视网膜的小双极轴突末端,靠近相关小神经节细胞的带状突触。
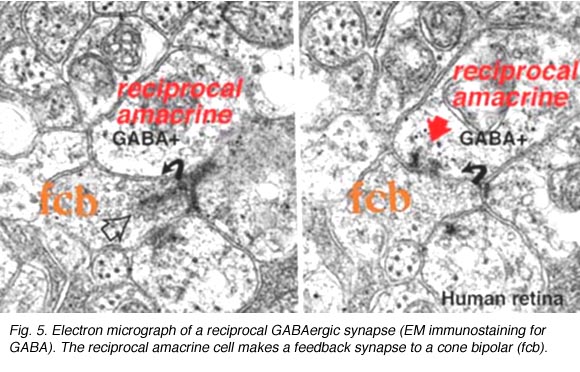
在这些情况下,反馈(互易)突触可能与这些细胞反应特征的阶段性或短暂成分有关。在输出到神经节细胞之前,它们可以通过反馈到双极细胞,使双极细胞缓慢的电位反应变得锋利和加快。
3.Interplexiform细胞。
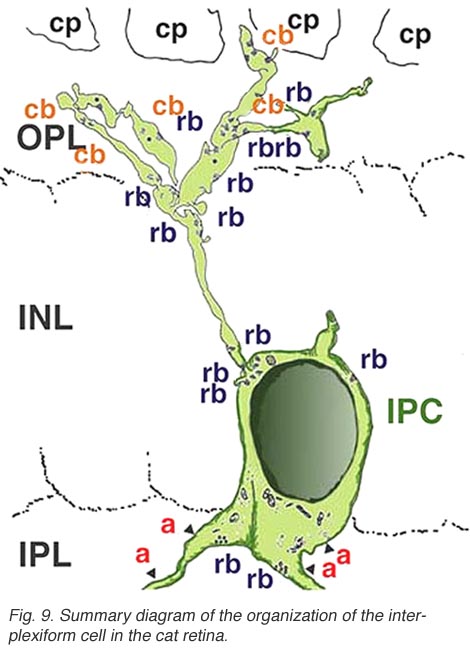
几乎所有被研究的物种,包括人类在内,视网膜中的神经元通过接受IPL的突触输入并在OPL的神经元上产生突触输出,将两个丛状层连接起来。它被称为丛状细胞IPC.Gallego(1971)首先描述了猫视网膜中的互丛细胞,Dowling和合著者(Dowling和Ehinger, 1975)在金鱼视网膜中进行了广泛研究,现在也在猫和人类视网膜中进行了研究(图6)(Kolb和West, 1977;林伯格和费雪,1986年;Kolb等人,1992)。在猫和人的这种互丛状视网膜中,细胞是中场的,在内丛层呈分支状三层。无论是从细胞体还是从IPL第1层的树突中,突起通过内核层上升到外丛状层。在那里他们跑了一小段距离(图6和图7)。
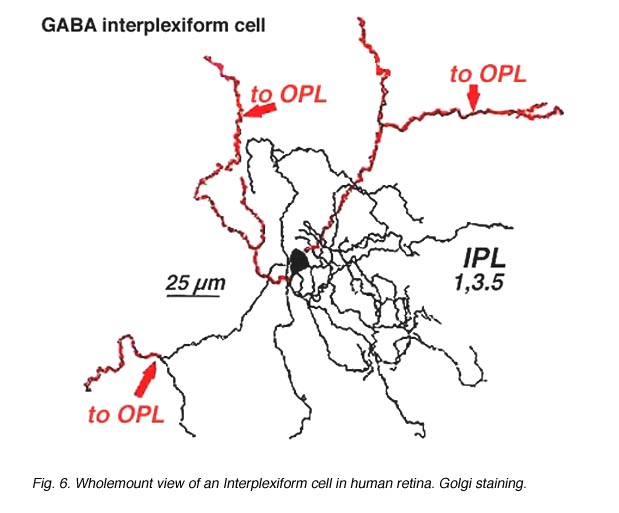 图6所示。人视网膜内丛状细胞的全景图 |
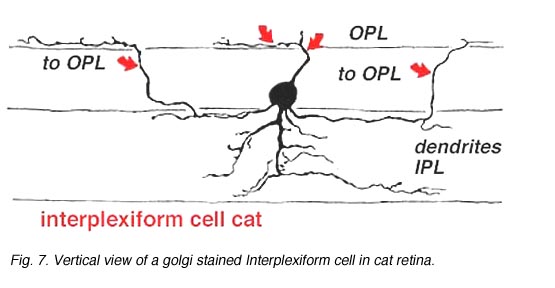 图7所示。人视网膜高尔基染色间丛状细胞的垂直视图 |
电子显微镜已经在猫和人的视网膜上对这种类型的IPC进行了例子。它们被证明从未知类型的无分泌细胞中获得输入,并在杆状和锥状双极轴突上产生一些突触(Kolb和West 1977;Nakamura et al., 1980)及其内部丛状层分支。IPCs细胞的过程通过内核层进入外丛状层,在杆状和锥状双极细胞体和顶端树突上产生大量常规突触(图8)。图9是由一系列EM图像重建的人类视网膜丛状细胞,并总结了这些细胞类型的连通性(Kolb and West, 1977)。
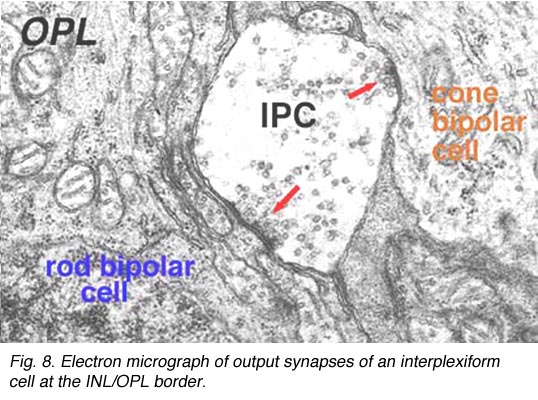
在猫和人身上,丛状细胞都有接触锥蒂的突起,但在这一部位形成相当非特化的连接,所以我们一直不愿意称这些为突触。刚刚在人和猫身上描述的互丛状细胞是已知的gaba能细胞。这与鱼视网膜的互丛细胞有很大的不同,在那里已知它是多巴胺能细胞(Ehinger等人,1969年)。在鱼的视网膜中,已知多巴胺能丛状细胞的下视区主要在水平细胞上有突触。因此,与人类的gaba能互丛细胞相比,鱼的互丛细胞似乎在视网膜中发挥了相当不同的作用。
4.哺乳动物的多巴胺能细胞。
在人和猫的视网膜中都存在一种叫做A18的多巴胺能无分泌细胞(图10),我们认为它参与了IPL的杆状系统通路,特别是杆状无分泌AII细胞(见前几章)。它们是大细胞体的无毛分泌细胞,具有长而多分枝的树突,主要局限于IPL第1层的丛状丛,位于无毛分泌细胞体下方(图10)。然而,有时可以看到这些A18细胞的突起从它们的细胞体或内丛状层第1层的树突丛上升到外丛状层。在这些层中,多巴胺能过程可能是人类视网膜水平细胞的突触前过程(Frederick et al., 1982)。然而,在猫的视网膜中,我们已经看到这些上升的多巴胺能过程(以其Toh免疫反应性的含量为标志)是传统的伽马能IPC的突触前过程(图11),即上述细胞(Kolb等人,1990,1991)。最近的证据表明,哺乳动物视网膜中的多巴胺能细胞是视网膜中含有黑视素的M1神经节细胞的突触前和突触后(Vuglar et al., 2007;张等,2008;McMahon等人,2010)。因此,多巴胺能无分泌可能从大脑的视交叉上核(SCN)接收离心输入,在那里昼夜节律是从含有黑视素的神经节细胞开始的。(参见黑视素神经节细胞章节)。 The dopamine cell in retina seems to be involved in feed-back loops from the SCN (through melanopsin cells) and within the retina modulating AII amacrine cell gap junctions, and from inner plexiform layer to outer plexiform layer influencing gap junctions between horizontal cells and even between photoreceptors.
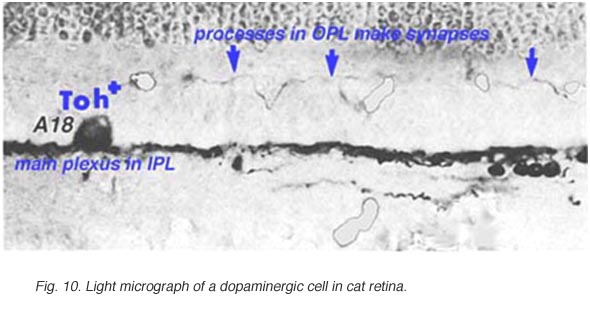 图10所示。多巴胺能细胞的光学显微镜图 |
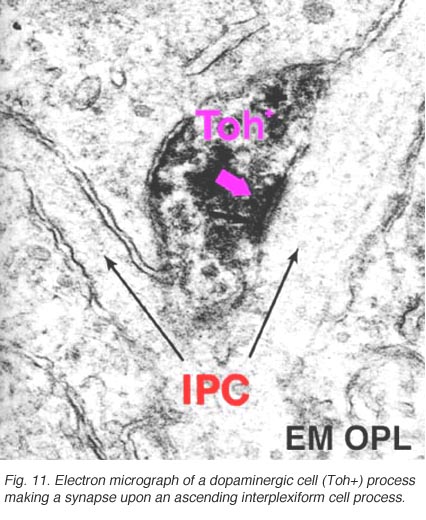 图11所示。多巴胺能细胞的电子显微镜图 |
5.哺乳动物视网膜中的离心纤维。
离心纤维产生于脑核并返回视网膜在哺乳动物中已被零星发现,但关于它们可能起源于哪个脑中心的细节和知识甚少。离心纤维在鸟类视网膜中特别发达(Cajal, 1892;Maturana和Frenck, 1965;奥格登,1968)。Polyak(1941)和后来Honrubia和Elliot(1970)分别在猴子视网膜中描述了一些离心纤维。根据Polyak(1941)的说法,猴子的离心纤维有静脉曲张,球茎末端在靠近无分泌细胞体的内丛状层结束。在人类视网膜中,它们穿过神经纤维层进入内核层后消失(Honrubia和Elliot, 1968年)(见下文)。
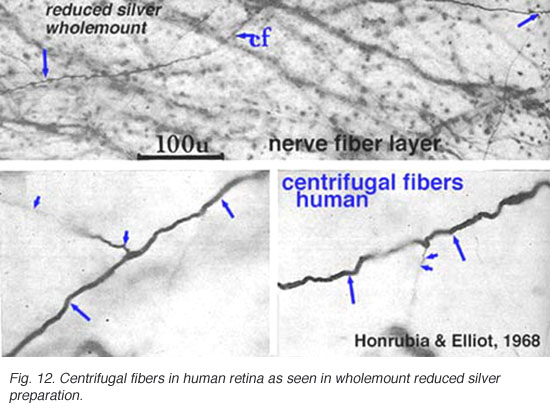 图12所示。人类视网膜中的离心纤维 |
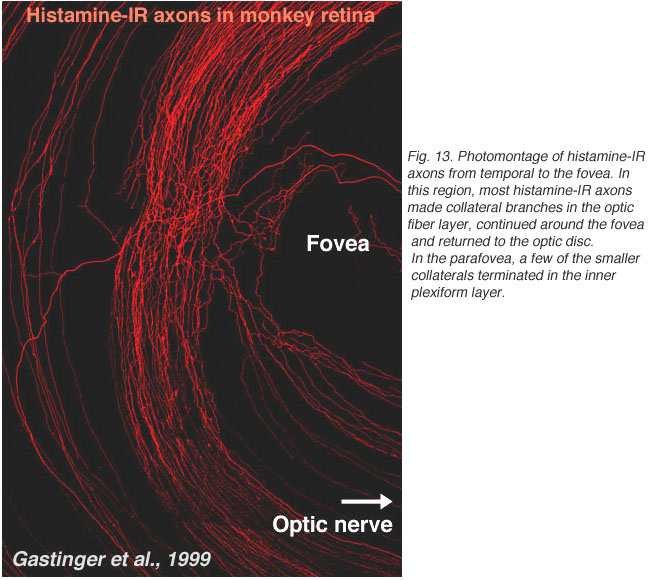 图13所示。从颞区到中央凹的组胺免疫反应轴突的蒙太奇。在这一区域,大部分组胺- ir轴突在光纤层形成侧枝,沿中央凹延伸并返回视盘。一个大轴突穿过这个密集的侧枝网络,最终形成一个垂直突出的分支到内丛状层。在旁凹区,少数较小的侧支终止于内丛状层,其余终止于光纤层的小肿胀 |
最近,人们发现,产生于下丘脑的离心轴突投射到大脑的不同部位,包括猕猴的视网膜。有趣的是,这些轴突含有组胺(Gastinger et al., 1999)。组胺免疫反应轴突在颞视网膜中央凹远侧的一个宽圆形带中从视神经头出发,似乎返回视盘(见图13)。这些轴突中的许多分支广泛,从神经纤维层延伸到内丛状层。有时组胺能轴突与视网膜血管有关。这将是非常有趣的发现,这些含有组胺的离心纤维是否与多巴胺无分泌或视网膜的其他细胞相互作用,就像其他物种的离心纤维一样。最近有证据表明,小鼠视网膜中的多巴胺能无分泌素对H1组胺受体发出的信号有反应(Frazao et al., 2011)。
在鱼视网膜中,我们知道离心纤维使用激素FMRFamide和黄体生成素释放激素作为神经递质(Stell, 1985)。它们在鱼体内直接与多巴胺互丛状细胞的细胞体相连(Zucker and Dowling, 1987)。在海龟的视网膜中,只有3 -6个对肽会面脑啡肽有免疫反应的传出纤维进入视网膜并向上传递到无分泌细胞区。这些纤维起源于尾端中脑,但它们在视网膜中的功能和作用仍不清楚(Weiler, 1985)。
6.参考文献
Baylor DA, Fuortes MGF, O 'Bryan PM。海龟视网膜上视锥细胞的感受区。杂志。1971;214:265 - 294。[PubMed] [免费全文在PMC]
卡哈尔老(1892)。在:Thorpe SA, Glickstein M,翻译。视网膜的结构斯普林菲尔德(伊利诺伊州):托马斯,1972年。
金鱼和Cebus猴视网膜含胺互丛细胞的突触组织。Science.1975; 188:270 - 273。[PubMed]
einger B, Falck B, lates AM。心侧链突中的肾上腺素能神经元。米克罗斯克·阿纳特。1969;97:285-297。[PubMed]
Frederick JM, Rayborn ME, Laties AM, Lam DM-K, Hollyfield JG。人类视网膜中的多巴胺能神经元。中华神经科学杂志1982;210:65-79。[PubMed]
加乐高。建筑学报。1971;31:299-304。
加斯廷格·MJ,奥布赖恩·JJ,拉森JNJ,马尔沙克DW。猕猴视网膜中的组胺免疫反应性轴突。眼科科学。1999;40:487-495 .[PubMed]
Honrubia FM, Elliott JH。视网膜的传出神经支配。一、人类视网膜的形态学研究。拱Ophthal。1968;80:98 - 103。[PubMed]
Honrubia FM, Elliott JH。视网膜的传出神经支配。2猴子视网膜的形态学研究。投资。Ophthal。1970;9:971 - 976。[PubMed]
Kolb H,西RW。猫视网膜内丛状细胞的突触连接。J Neurocytol。1977;6:155 - 170。[PubMed]
王红红,王红红。猫视网膜中多巴胺能无分泌细胞的突触组织。J Neurocytol。1990;19:343 - 366。PubMed]
GABA和甘氨酸的包埋后免疫细胞化学揭示了猫视网膜多巴胺能无分泌细胞的突触关系。中华神经科学杂志1991;[PubMed]
高尔基体对人类视网膜神经元的研究。中华神经科学杂志1992;318:147-187。[PubMed]
Linberg KA, Fisher SK.视网膜中水平细胞轴突终末是突触前的超微结构证据。中华神经科学杂志1988;268:281-297 .[PubMed]
鸽子视网膜中离心纤维的突触连接。科学。1965;150:359 - 361。[PubMed]
Nakamura Y, McGuire BA, Sterling P.猫视网膜中的丛状细胞:通过摄取[γ]-[3.H]氨基丁酸及其序列重建。美国国家科学院1980;77:658-661。[PubMed] [免费全文在PMC]
奥格登TE。视网膜传出纤维的功能。在:抑制性神经元机制的结构和功能。纽约:佩加蒙出版社;1968.p . 89 - 109。
Polyak SL,视网膜。芝加哥:芝加哥大学出版社,1941
民谣钢弦工作。推测肽递质,无分泌细胞多样性和功能在内丛状层。作者:Gallego A, Gouras G,编辑。视网膜的神经回路:卡哈尔的纪念。纽约:爱思唯尔;1985.p . 171 - 187。
龟视网膜的传入和传出肽反应通路。作者:Gallego A, Gouras P,编辑。视网膜的神经回路:卡哈尔的纪念。纽约:爱思唯尔;1985.p . 245 - 256。
朱克CL,道林JE。硬骨鱼视网膜多巴胺能丛状细胞上的离心纤维突触。大自然。1987;330:166 - 168。[PubMed]
最后更新:2011年7月。