海尔格科尔布
1.一般特征。
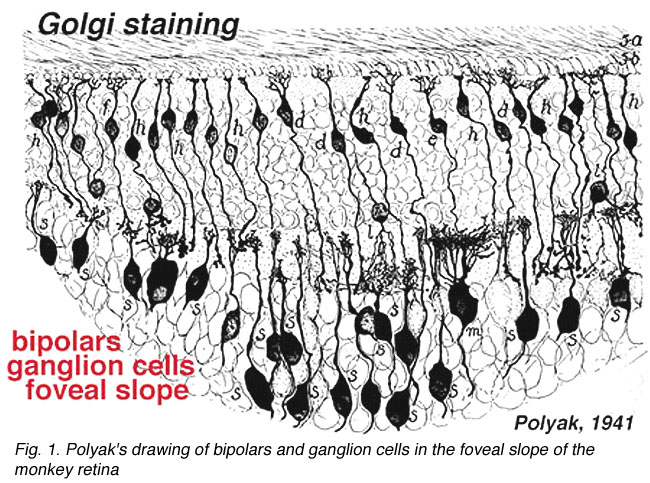
人和猴视网膜中央凹的特殊锥体通道收敛性最低,视觉系统分辨能力最强。beplay体育公司这是通过使连接尽可能“私密”并在所谓的小路径中将它们缩小为一对一的关系来实现的。图1显示了Polyak的书(!941)中的一幅古老的画,显示了最窄的视野双极细胞和中央凹的神经节细胞。
图1。Polyak绘制的猴子视网膜中央凹斜坡的双极细胞和神经节细胞。插图来自1941年的Polyak。
小通路由小双极细胞和小神经节细胞组成,小神经节细胞投射到大脑外侧膝状核的单个细小细胞层细胞上。因为需要高灵敏度的小通路也被组织成像弥漫性锥体通路一样的ON-和OFF-center通道,以最大化对比度,这意味着每个中央凹锥体将有双小通路。这两个小双极将是ON-center型和OFF-center型,分别连接ON-center和OFF-center侏儒神经节细胞。因为小双极/神经节细胞电路只使用单一的锥体,而且每个锥体具有不同的光谱类型,由短(s -锥体/蓝色)、中(m -锥体/绿色)或长(l -锥体/红色)波长最大程度激发,它们必须携带一个波长的灵敏度信息。因此,特别是与M-和L-视锥有关的小系统也被认为是将L-或M-信号传送到大脑,在那里进行进一步的处理,使我们能够辨别颜色视觉中的红色和绿色。
s锥系统与M锥和l锥系统从锥体本身的初始水平到通过视网膜进入大脑的回路都有很大的不同。我们不认为s锥通路是一个小通路(参见s锥通路后面的章节)。所以这一章专门讲M-和l -小通路我们会看到,它们通过视网膜传递分辨率和颜色信息到大脑。
2.视觉敏锐度始于锥体间距和小电路。
视敏度是衡量我们辨别最细微事物能力的一种方法:可以是辨别两条平行线(游标视敏度),也可以是辨别两个小点或星星(点敏度)。灵长类动物和人类的视觉系统能够达到1分弧度或beplay体育公司60循环/度的视角的敏锐度。一个角度被认为可以覆盖大约280-300微米的视网膜距离(Drasdo和Fowler, 1974)。视网膜上视锥感光细胞最紧密地排列成或多或少的六角形排列的区域,当然是中央凹的中心,大约有一个度(图2)。因此,1分弧度的视觉分辨大约是中央凹中镶嵌视锥的中心到中心的间距(3µm)(图3)。
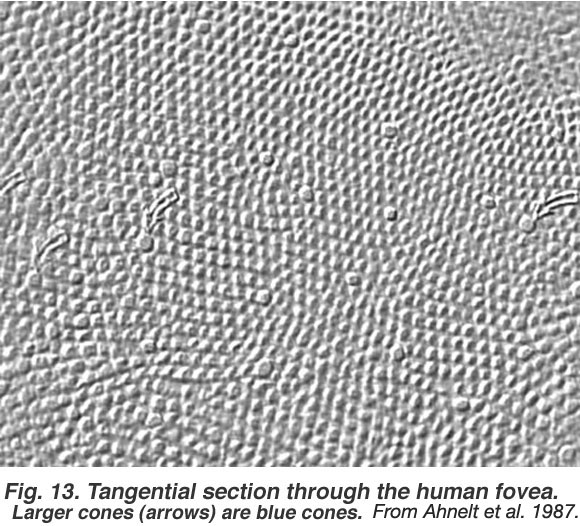 图2。穿过人的中央凹的切向切片。较大的球果(箭头)为蓝色球果。插图来自Ahnelt等人,1987年。 |
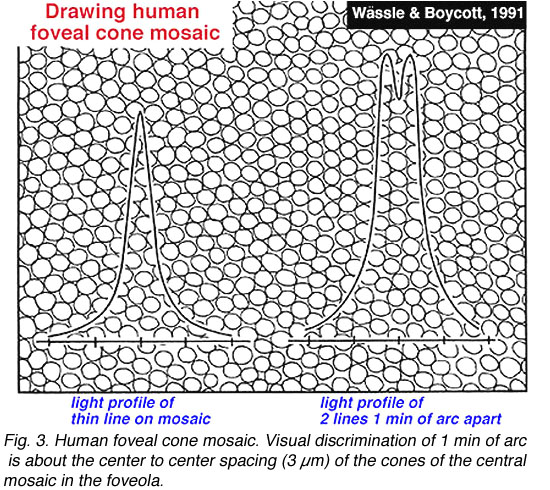 图3。人类中心凹锥体镶嵌。1分圆弧的视觉分辨大约是凹中央镶嵌锥体的中心到中心间距(3 μm)。插图来自Wässle和Boycott, 1991年 |
如果最小中心距(一个)为3 μ m,则光栅分辨率极限即奈奎斯特极限为一个√3,约1分弧。有人可能会问,为什么光栅的分辨率不优于1分弧和大小,甚至小于每个锥体的直径,即。一个或<一个.对于这些较窄的光栅,如果线分离小于,填充锥体的空间干涉显然会引起云纹效应或混叠一个使用√3,如下所示(Wässle和Boycott, 1991)(图4)。这些干涉图样的重复长度取决于光栅与锥镶嵌的角度,因此无法确定光栅的波长。事实证明,在正常情况下,眼睛的光学不能清晰地成像超过60循环/度的光栅(Cambell和Gubisch, 1966)。在空间采样中,眼睛的光学模糊最小化了云纹效应(图4)。
图像也被眼睛的光学模糊,投射到大约10个视锥细胞上(图4)。然后,图像的横向移动可以在这10个视锥细胞和它们的小神经元链上进行比较,以供稍后在大脑中进行计算。这使得更好的辨别(分辨)成为可能,因为邻近的锥系统(即路径上的周围环境)可能产生对抗,并增加了几个小型-单锥起源路径之间的对比和辨别(Gouras, 1992)。

我们知道三种类型的锥体组成锥体中心凹镶嵌,但我们一直很好奇这三种不同光谱类型的锥体是如何排列在六边形镶嵌中。s锥(蓝色锥)很容易从l锥(红色锥)和m锥(绿色锥)中分辨出一些独特的形态特征(参见s锥路径一章)。事实上,在图2的中心凹镶嵌全景图中,s锥被视为破坏六边形镶嵌的较大轮廓(箭头指向s锥)。在圆锥花叶的中心凹区,8-12%的圆锥形成s -圆锥(Ahnelt et al., 1987)。中央凹的中心几乎没有s锥。所以剩下的球果以某种方式被分为L型和m型球果群体。在人类的中央凹,心理物理测量表明红色视锥细胞的数量是绿色视锥细胞的2:1 (Cicerone和Nerger, 1989)。然而,通过显微分光光度法直接测量猴子中央凹小块锥体中的所有锥体,发现红色和绿色锥体的出现比例大致相同(图5)(Mollon和Bowmaker, 1992)。Roorda和Williams(1999)通过一种复杂的自适应光学技术来提高空间分辨率,直接测量了活体人眼的中心凹锥的光谱灵敏度。他们发现人类在红色球果和绿色球果的比例上差异很大:有些人的比例几乎相等,而另一些人的红色球果比例更高,达到了2个红色球果比1个绿色球果。 The red and green cones lie randomly in the mosaic meaning that clusters of cones of the same spectral type will occur together as illustrated in the figure (Fig. 5, below) from Mollon and Bowmaker’s paper. An interesting point can be deduced from these findings. It does appear possible then, that a multiheaded midget or diffuse cone bipolar with a small dendritic tree, could contact all the same spectral type of cones in the primate fovea.

图5。猕猴锥体中心凹斑块三种锥体类型的吸收光谱测定。这些锥体斑块的光谱组成显示。插图来自Mollon和Bowmaker, 1992年
3.锥体到小双极细胞连接。

1941年,Polyak(图6)已经在灵长类动物视网膜上描述了两种类型的小神经节细胞。他从高尔基染色猴视网膜的垂直切片上研究并绘制了神经节细胞,并注意到,尤其是小神经节细胞(尽管大小阳伞神经节细胞也是如此),其树突在IPL的两个层次之一分支:靠近神经节细胞体(我们现在称为亚层)的区域b)或在靠近无分泌细胞体(我们现在知道的亚膜区域)的神经泌中一个).
图6。Stephen L. Polyak(1889-1955)是灵长类动物眼睛解剖学和神经组织方面的主要权威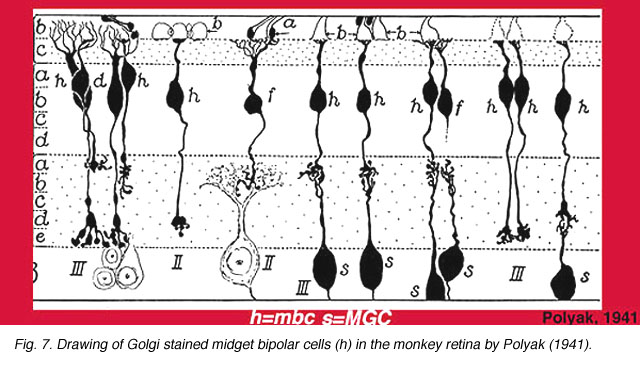
他还绘制并评论了这样一个事实,即小双极细胞有长轴突和短轴突的变种,以适应两个小神经节细胞变种(图7)(Polyak, 1941)。
两种类型的小双极细胞之间的重要区别直到电子显微镜研究能够显示它们与锥椎弓根的突触接触的差异(Kolb, 1970)。
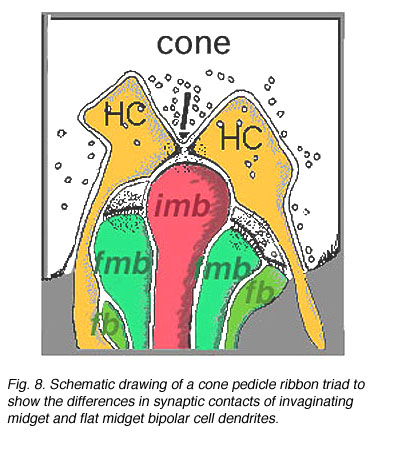
图8。锥体蒂带状三联体示意图,以显示凹陷小体和扁平小体双极细胞树突突触接触的差异。
因此,内陷的小双极细胞(imb)被发现与锥体蒂的中央单元连接,与锥体蒂带的内陷突触连接(图8和9)。扁平的小双极细胞在宽裂的基底连接(fmb)与锥体蒂连接,最常位于imb的内陷树突的两侧(图8和10)。
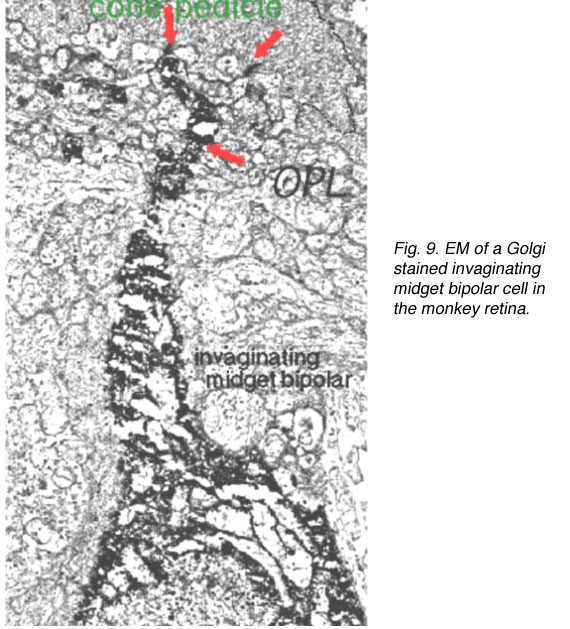 图9。猴子视网膜高尔基染色内陷小双极细胞的EM图。OPL,外丛状层。 |
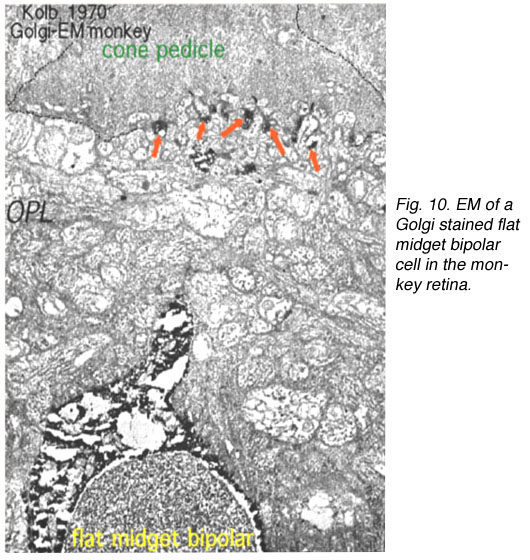 图10。猴子视网膜高尔基染色扁平小双极细胞的EM图。OPL,外丛状层。这幅插图来自科尔布,1970年 |
4.小型神经节细胞。
观察高尔基染色的整个灵长类动物视网膜特别有助于揭示与空间和颜色视觉有关的三种神经节细胞类型之间的差异:即侏儒神经节细胞、蓝色/黄色神经节细胞和阳伞神经节细胞(Polyak, 1941;Kolb等,1992;达西和李,1994年)。小神经节细胞被认为是高敏锐度细胞,也携带红色或绿色的特定信号。它们投射到外侧膝状核的细小细胞层,因此称为P细胞(图11)(Shapley and Perry, 1986)。小神经节细胞有高分支的种类,在生理上可能是偏中心的,而低分支的类型在生理上是on中心的。

图11。人视网膜遮阳节(M)和小节(P)细胞的高尔基染色。所有图像比例尺为10 μm。插图来自Kolb, Lindberg和Fisher, 1992年
点击这里查看人类视网膜中两个高尔基染色P细胞或小神经节细胞的聚焦系列(Quicktime电影)。一对高尔基染色的人小神经节细胞,包括ON型和OFF型,显示了视网膜平贴视图中不同的树突分支水平。
图11显示了两个小神经节细胞与周围人视网膜的阳伞神经节细胞(M细胞)的比较。一个小的神经节细胞有直径18微米的树突树,另一个有直径35微米的更大的树突场。上面的影片显示,树突场较小的小神经节细胞与树突场较大的细胞在不同的水平上聚焦。前小神经节细胞在IPL中分支高,因此在生理上可能是OFF中心型。树突状树较大的分支较低的小神经节细胞可能是ON中心型。为什么OFF中心型的树突树比ON中心型的更小,目前仍然是一个谜,但一个可能的含义是,小型OFF系统的灵敏度高于ON系统(Kolb和Marshak, 2001)。
小双极和小神经节细胞之间的连接一直被认为是“私人的”,即一对一的。由于双极细胞的轴突末端和小神经节细胞的树突杆的大小和分支水平相似,特别是在中心凹区,我们认为轴突末端和树突场重叠,并且突触,每个神经节细胞没有一个以上的双极轴突收敛的空间。然而,周围视网膜的情况可能不同,那里的小神经节细胞有更大的树突场。如图12所示,明显多于一个小双极轴突可以与接收周边小神经节细胞的树突树相匹配。

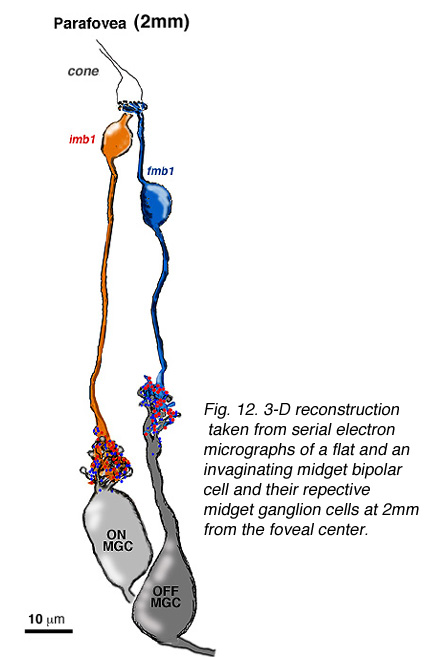
图12。小双极细胞(imb, fmb)在离中心凹更远的地方有更大的轴突末端(视网膜偏心4.5mm,而视网膜偏心12 mm)。小神经节细胞(MGC)有更大的树突场(这里看到8毫米),因为他们增加偏心。
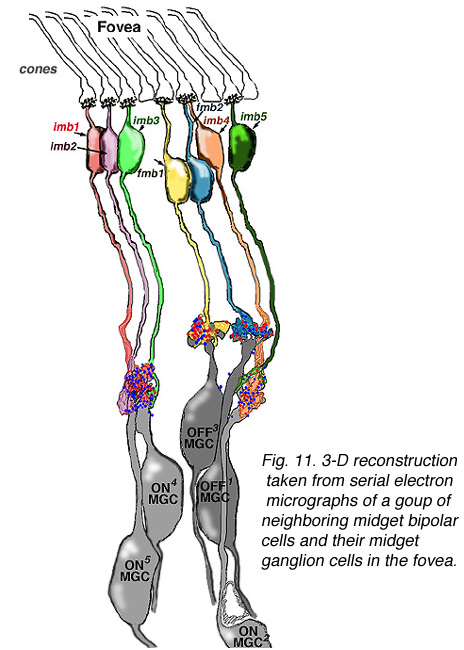
通过电子显微镜研究和重建距离中心凹约2毫米的旁中心凹连续切片的小神经节细胞及其输入的小双极细胞轴突,证实了这种一对一的关系(图14)(Kolb和DeKorver, 1991)。最近,我们还对中心凹侏儒双极轴突及其与中心凹侏儒神经节细胞树突的联系进行了一系列切片电镜研究(图13)(Kolb和Marshak, 2003)。
 图13。从一组相邻的小双极细胞及其突触后小神经节细胞的连续电子显微图中获得的人中央凹的三维重建。 |
 图14。从距离人视网膜中心2mm处的扁平和凹陷的小双极细胞及其各自的小神经节细胞的连续电子显微图中获取的三维重建。 |
{kind=link}
{kind=link}
各自的小双极细胞几乎完全是单个小神经节细胞上的突触(图14),除了在非常中央的中央凹,由于神经元和神经节细胞的拥挤,一些小双极突触与邻近的小神经节细胞共享(图13)。因为我们知道(Nelson et al., 1978)神经节细胞在椎板下分支一个会偏离中心,而这些分支在亚椎间b我们可以确定靠近无分泌细胞层分支的小神经节细胞是OFF中心的,而靠近神经节细胞层分支的小神经节细胞是ON中心的。这种开和关的灵长类小神经节细胞组织已经被Dacey和合作者(2000)证实。因此,在中心凹区域和视网膜中央边缘(距离中心凹中心约4毫米),人类中央凹的小通路以以下方式组织:1个锥到2个小双极细胞(ON-和off -中心双极类型)到2个小神经节细胞(ON-和off -中心神经节细胞类型)。
我们发现比较难说的是在小双极和小神经节细胞链中连接的是哪种光谱类型的锥体。M型和l型锥在解剖学上难以区分。我们可以在不寻常的形态特征上识别s锥(蓝色)蒂(Ahnelt et al., 1987, 1990;Kolb et al., 1997)(参见s锥通路章节),M锥和l锥看起来是一样的。有证据表明M-和l -连接的小双极细胞有不同数量的与轴突相关的突触带,因此在中心凹有不同数量的输出到其小神经节细胞的位点(Calkins et al., 1994):但轴突末端/神经节细胞对与M-或l -锥通路相关仍是一个有争议的问题。
在中央凹外4 - 5毫米处,在近周围,小双极细胞变成2和3头分别连接2和3个锥细胞(Polyak, 1941;Kolb等,1992;博伊特和霍普金斯,1991)。小神经节细胞遍布近周和中周视网膜(Polyak, 1941;Rodieck等,1985;达西和彼得森,1992年;达西,1993;Kolb et al., 1992)和它们的树突树的大小增加,但通常由多个树突域组成。小神经节细胞的树突状树的结构域越近,树突状树的结构域越多。所以看起来很可能不止一个小双极细胞会在每个小神经节细胞上产生突触,可能每个双极细胞占据树突树的一个区域。
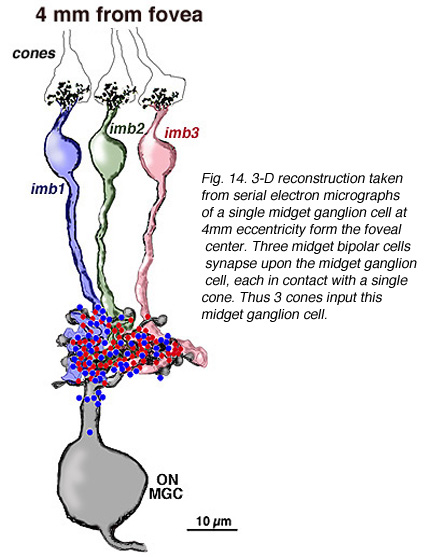
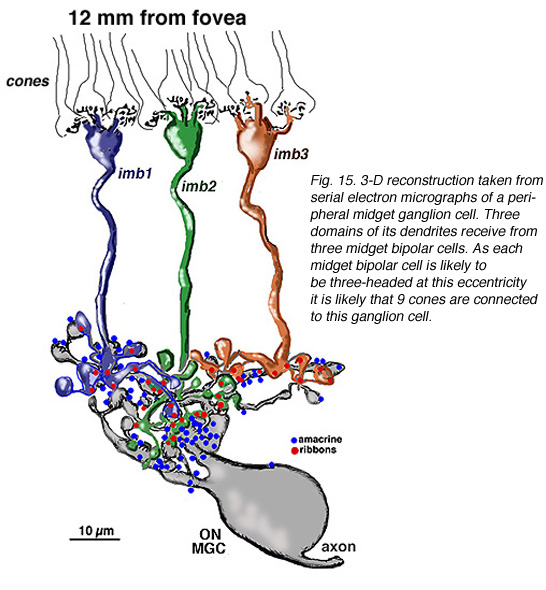
在中心区域边缘(即距中心凹4毫米处)和周围视网膜(距中心凹12毫米处)的小神经节细胞的轴突和树突的连续切片电镜和重建表明,在周围视网膜中,锥细胞对小双极细胞的多重输入和小双极细胞对小神经节细胞的多重输入的预测是正确的(图15和16)。我们的估计(Kolb和Marshak, 2003)是3个侏儒双极细胞在一个离中心凹4毫米的神经节细胞上形成突触(图15)(Kolb和Marshak, 2003)。在远周视网膜上,我们估计一个小神经节上至少有3个多头小双极细胞的突触(图16)。
 图15所示。猕猴视网膜小神经节细胞在距中心凹偏心4mm处的连续电子显微图上的三维重建。三个小双极细胞在小神经节细胞上突触,每一个都与单个锥体相连。因此有3个视锥细胞输入这个小神经节细胞 |
 图16所示。从周边小神经节细胞的连续电子显微图中获得的三维重建。其树突的三个区域来自三个小双极细胞。由于每个小双极细胞在这个偏心率下很可能是三个头,因此很可能有9个锥细胞连接到这个神经节细胞 |
{kind=link}
{kind=link}
在中央凹,一个单一的锥体与它的小双极和小神经节细胞系统连接,通路将必然携带一个单一的颜色感受野中心。在小脑中央凹以外,即小神经通路为2通道和3通道和/或许多小双极患者有输入,将小神经节细胞的中心可能是锥体颜色类型的混合。尽管红色和绿色锥体马赛克和多个小双极输入的斑块和随机性质,但小神经节细胞是否保留对一个光谱类锥体的主导承诺,从而形成一个颜色对手中心环绕组织?这实际上是两种生理学(Martin et al., 2001;Reid和Shapley, 1992)和Chichilnisky团队最新的功能连通性报告(图17)(Field等人,2010)。
图17显示了使用多电极记录和高分辨率刺激这些神经节细胞感受野中包含的单锥类型的小神经节细胞测量结果。虽然红色和绿色的视锥细胞都输入细胞,但单一光谱类型的视锥,即红色或绿色类型的视锥,主导着接受野中心(图17,白色轮廓和白线)。

5.人类视网膜中红色和绿色对抗的基础电路。
已知猴子视网膜(可能也是人类视网膜)的小神经节细胞对光线的反应与周围的相对色组织(Gouras, 1968, Gouras, 1991;Dacey和Packer, 2003,回顾)(图18)。也就是说,视网膜中央的小神经节细胞,从电生理上记录,有最小的感受野,并组织为l锥ON或OFF中心和m锥ON或OFF中心。每一种小神经节细胞类型都有一个较大的相反极性和相反颜色的包围。小神经节细胞的周围,正如Chichilnisky的团队优雅地展示的(Field et al., 2010),周围由许多红色和绿色锥输入组成,表明对周围非特异性的,但混合的光谱输入(图17,e)到g)(黑线)。
 图18。猴子视网膜上记录的对色单元。插图来自Gouras, 1968(28)。 |
{kind=link}
神经节细胞的阳伞型是视网膜中记录的神经节细胞的非颜色对位相型(Gouras, 1968),也被称为M细胞,投射到外侧膝状核上(Shapley and Perry, 1986;威塞尔和胡贝尔,1966年;Gouras, 1992年的回顾)。视网膜和膝状核的细小细胞层也记录了一个较小的场、强韧且不清楚颜色的对位神经节细胞类型(图18)(Gouras, 1992)。这些是否是少数的中央凹外小神经节细胞,它们不再像Field等人的研究(2010)中的一些细胞那样具有颜色特异性,还有待观察。
在小神经节细胞中记录的对抗性颜色包围(Gouras, 1968)提供颜色和空间上的对抗性。现在看来,所有灵长类锥体双极性类型都具有中心/环绕结构(图19)(Dacey et al., 2000;Field等人,2010)。这包括小双极细胞(图19)(Dacey等人,200)。这种对抗性的周围组织从何而来?

在亚哺乳动物中,水平细胞被认为有助于双极细胞的环绕反应,特别是在具有良好色觉的动物中。例如,在鱼和海龟中,水平细胞已被证明是颜色对手(Stell和Lightfoot, 1975;参见Kamermans和Spekreijse, 1995或Kolb和Lipetz, 1991的评论)。灵长类动物视网膜的情况则不同,在猴子水平细胞的细胞内记录中,尽管有光谱选择性连通性的解剖学发现(Ahnelt和Kolb, 1994;Dacey et al. 1996)。
然而,最近在猕猴身上进行的优雅实验表明,红色和绿色视锥细胞从水平细胞(可能是HI细胞)处接受拮抗包围反应(Verweij et al., 2003)。类似地,根据Packer和合作者(2010)的说法,灵长类动物(蓝色)s锥细胞明显有一个相对的黄色包围,似乎来自L-和m -锥细胞驱动的水平细胞。来自同一组的最新证据(Crook等人,2011年)相当明确地表明,水平细胞反馈可以解释灵长类动物视网膜中红色和绿色小神经节细胞周围的竞争,表明小双极细胞在本质上已经是颜色对手。在小双极细胞和小神经节细胞中,似乎没有任何视网膜内部(无分泌细胞)输入来组织周围。存在的可能性是,一些大场扩散锥双极类型,没有光谱对抗,并且其周围一直比小型双极细胞更大,可能涉及无分泌细胞含量(泰勒,1999;Dacey et al., 2000)。

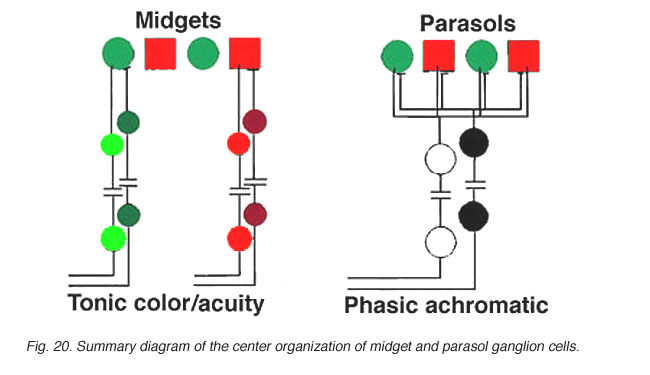
图20。灵长类动物视网膜上的小通路,其中心和周围(红色/绿色)颜色相对,中心和周围(ON(+)或OFF(-)颜色相对。
我们认为L-和m -小神经节细胞反应的组织方式如图20所示。一个l -锥体会接触两个l -锥体侏儒双极子,通过它们,两个l -锥体侏儒神经节细胞。例如,l -锥体on -中心小双极细胞会接触到一个l -锥体位于前下丘脑的内陷突触处,以及一个l -锥体位于on -中心的小神经节细胞bIPL。同样的l -锥体在基底连接处也会被一个off -中心的小双极细胞接触,而双极细胞反过来又会接触到一个off -中心的小神经节细胞一个(图20,左)。这样就产生了l锥ON和l锥off中心感受野神经节细胞类型(图20,左)。m -视锥细胞将以类似的方式连接到两个侏儒双极细胞(ON和OFF型)和两个侏儒神经节细胞,它们也将是ON和OFF中心感受野类型(图20,右)。
HI、HII和HIII三种水平细胞均能使m锥胞体向l锥中心双极细胞转变,l锥胞体向m锥中心双极细胞转变。然而,HII细胞可能更多地参与蓝/黄两极细胞通路的光谱拮抗场(见网络视觉s势和水平单元).
点击此处观看人类视网膜中侏儒通道的动画(Quicktime电影)
6.M和蓝色/黄色神经节细胞。
灵长类动物视网膜上的大视野阳伞细胞被认为传递有关低空间灵敏度、运动和亮度信息的消色差信息(Merigan, 1989;Merigan和Maunsell, 1990)。像小神经节细胞一样,阳伞细胞以ON-和off -中心的形式出现,并投射到外侧膝状核的巨细胞层(图21):因此它们也被称为M细胞(Shapley和Perry, 1986;卡普兰和沙普利,1986)。阳伞神经节细胞对其感受野生理也有环绕机制,这与小神经节细胞一样,被认为主要来自于外丛状层的水平细胞反馈机制(McMahon et al.,(2004)。蓝黄色神经节细胞(图21)是一种双层类型,被认为是由蓝色特异性双极细胞和其他小型或弥漫型细胞驱动的,从而产生黄色反应(Dacey和Lee, 1994)。它们投射到膝状核的一个特殊层,称为koniocellular layer或K layer (Martin et al., 1997;Roy等人,2009)(图21)。我们将在另一章讨论短波敏系统(Webvision: S-cone通路)所以我就不多说这些神经节细胞了。

图21。灵长类神经节细胞投射到膝状外侧核和上丘的图示。上面的插图B/Y来自Dacey和Lee, 1994(11)。LGN的插图来自Shapley和Perry, 1986年
7.参考文献
在人类视网膜中发现一种可能对蓝色敏感的视锥光感受器亚型。J Comp Neurol.1987; 255:18-34。[PubMed]
王晓燕,王晓燕,王晓燕,等。人眼和灵长类动物视网膜中蓝色敏感视锥细胞蒂的鉴定。中华神经科学杂志1990;293:39-53 .[PubMed]
视网膜的水平细胞和锥感光体:光谱连通性的高尔基电子显微镜研究。J Comp Neurol.1994; 343:406 - 427。[PubMed]
抵制BB,霍普金斯JM。灵长类动物视网膜中的锥体双极细胞和锥体突触。Vis > 1991; 7:49-60。[PubMed]
猕猴中央凹的Calkins DJ、Schein SJ、Tsukamoto Y、Sterling p.m和L锥通过不同数量的兴奋性突触连接到小神经节细胞。大自然。1994;371:70 - 72。[PubMed]
Calkins DJ, Tsukamoto Y, Sterling P.灵长类动物视网膜中蓝黄色神经节细胞的微电路和马赛克。J > 1998; 18:3373 - 3385。PubMed]
Calkins DJ, Meszler LB, Hendry SHC。灵长类动物视网膜中多种神经节细胞表达II型钙调素依赖性蛋白激酶的亚基。社会神经科学文摘1997;23:729。
坎贝尔FW,古比什RW。人眼的光学质量。杂志。1966;186:558 - 578。[PubMed] [免费全文在PMC]
导游CM, Nerger JL。人中央凹长波长敏感锥体与中波长敏感锥体的相对数目。视觉Res.1989; 29:115 - 128。[PubMed]
人视网膜中小神经节细胞和阳伞神经节细胞的树突状场大小和形态。中国科学(d辑:自然科学版)1992;[PubMed] [免费全文在PMC]
人类视网膜中小神经节细胞的镶嵌。J > 1993; 13:5334 - 5355。[PubMed]
Dacey DM, Lee BB。灵长类动物视网膜上的“亮蓝”通路起源于一个独特的双层神经节细胞。大自然。1994;367:731 - 773。PubMed]
Dacey DM, Lee BB, Stafford DK, Pokorny J, Smith VC。灵长类动物视网膜的水平细胞:无光谱对抗性的锥特异性。Science.1996; 271:656 - 659。[PubMed]
ddacey DM, Packer OS, Diller L, Brainard D, Peterson B, Lee B.灵长类动物视网膜锥体双极细胞的中心环绕接受野结构。视觉Res.2000; 40:1801 - 1811。[PubMed]
Drasdo N, Fowler CW。广角原理图中视网膜图像的非线性投影。中华眼科杂志1974;58:709-714。[PubMed] [免费全文在PMC]
猴神经节细胞锥体机制的鉴定。杂志。1968;199:533 - 547。[PubMed] [免费全文在PMC]
色彩视觉的皮层前生理学。编辑者:古拉斯·P。视力和视觉障碍。第六卷:色彩的知觉。伦敦:麦克米伦出版社;1991.p . 163 - 178。
视网膜回路及其与诊断精神物理学和电生理学的相关性。Curr Opin Ophthal. 1992; 3:803-812。
Irvin GE, Casagrande VA, Norton TT。灵长类动物外侧膝状核中巨细胞、细小细胞和巨细胞中继细胞的中心/周围关系。Vis > 1993; 10:363 - 373。[PubMed]
王晓峰,王晓明。硬骨鱼视网膜中锥驱动水平细胞的光谱行为。Prog Ret Eye Res. 1995; 14:13 - 360。
Kaplan E, Shapley RM。灵长类动物的视网膜包含两种类型的神经节细胞,具有高和低的对比敏感度。中国科学(d辑:自然科学版)1986;[PubMed] [免费全文在PMC]
灵长类动物视网膜外丛状层的组织:高尔基细胞浸润的电子显微镜。费罗斯反式。r . Soc。B.1970; 258:261 - 283。
人视网膜旁凹小神经节细胞的电镜观察和序列切片重建。中华神经科学杂志1991;303:617-636。[PubMed]
Kolb H, Lipetz LE。(1991)。视网膜:脊椎动物视网膜上颜色视觉的解剖学基础编辑者:古拉斯·P。视力和视觉障碍。第六卷:色彩的知觉。伦敦:麦克米伦出版社;1991.128 - 145页。
高尔基体对人类视网膜神经元的研究。中华神经科学杂志1992;318:147-187。[PubMed]
柯柏H, Goede P, Roberts S, McDermott R, Gouras P.人类视网膜S锥蒂的独特形态及其对颜色处理的影响。中华神经科学杂志1997;386:443-460。[PubMed]
Kolb H, Marshak D.灵长类动物视网膜的小通路。医生角膜切削。2003;106:67 - 81。[PubMed]
Merigan WH。猕猴的彩色和无色视觉:P通路的作用。J > 1989; 9:776 - 783。[PubMed]
Merigan WH, Maunsell JH。巨细胞外侧膝状肌损伤后的猕猴视力。Vis > 1990; 5:347 - 352。[PubMed]
Mollon JD, Bowmaker JK。锥体:灵长类动物中央凹中锥体的空间排列大自然。1992;360:677 - 679。[PubMed]
Nelson R, Famiglietti EV, Kolb H.细胞内染色显示猫视网膜上和非中心神经节细胞有不同程度的分层。J Neurophysiol。1978;41:472 - 483。[PubMed]
Polyak SL,视网膜。芝加哥:芝加哥大学出版社,1941
Rodieck RW, Binmoeller KF, Dineen JT。人视网膜的阳伞和小神经节细胞。中华神经科学杂志1985;233:115-132。[PubMed]
Roorda A, Williams DR.活人眼睛中三个视锥类的排列。大自然。1999;397:520 - 522。[PubMed]
沙普利R,佩里VH。猫、猴视网膜神经节细胞及其视觉功能作用。趋势> 1986;9:229 - 235。
Stell WK, Lightfoot DO。金鱼视网膜中视锥细胞和水平细胞的颜色特异性相互联系。中华神经科学杂志。1975;159:473-501 .[PubMed]
Wässle H,抵制BB。哺乳动物视网膜的功能结构。杂志启71:447 1991;480年。[PubMed]
威塞尔TN,胡贝尔DH。恒河猴外侧膝状体的空间和颜色相互作用。J Neurophysiol。1966;29:1115 - 1156。PubMed]
最后更新:2012年6月。