斯图尔特Trenholm1以及高塔姆·b·阿瓦拉玛尼2
1加拿大蒙特利尔麦吉尔大学蒙特利尔神经学研究所助理教授:stuart.trenholm@mcgill.ca
2维多利亚大学生物系副教授,加拿大维多利亚:gautam@uvic.ca
摘要
在电子显微镜下,缝隙连接是两个细胞之间相对的膜上密集的淀粉区。小的示踪分子如神经生物素通过缝隙连接孔,标记相互连接的细胞。电信号通过这些连接,扩展了神经元数据收集的区域,修改了信号的动态特性,并同步脉冲的产生。在视网膜中,由多种神经元组成的电路将视觉环境中的信息转化为一系列经过高度处理的简化的脉冲序列,再传递到更高的视觉区域。视网膜信号处理依赖于特定细胞类型之间复杂的相互作用,这是由化学和电突触介导的。在该领域,在了解视网膜细胞类型的多样性(其中可能至少有100种独特类型)以及它们如何连接形成细胞类型特定的电路方面已经取得了巨大进展。在这里,我们概述了视网膜中的电耦合,并概述了间隙连接在健康和患病状态下处理信息时所起的不同作用。
简介
视网膜由大约100种不同的神经元细胞组成,它们位于特定的视网膜层,彼此连接形成复杂的微电路(1-5)。这些回路用于提取视觉世界的基本特征,这些特征通过不同类型的视网膜神经节细胞被解析成不同的视觉通道。这些差异处理的视觉信号随后从视网膜传递到大量的高级视觉区域(6,7)。有趣的是,视网膜中的微电路是通过化学突触和由间隙连接形成的双向电突触连接的。电突触和化学突触之间丰富的相互作用使电路以高度灵活和动态的方式运作。此外,由于间隙连接的强度可以在多种时间尺度上修改,它们可以在毫秒时间尺度上以及在白天和黑夜周期的过程中重新配置视网膜微电路。此外,间隙连接在一系列空间尺度上发挥作用,在亚细胞水平上同步活动,以及在视网膜的宽条带上。鉴于间隙连接在视网膜信号处理中的不同作用,电突触在疾病状态中也发挥许多作用也就不足为奇了。最后,尽管我们在理解缝隙连接在视网膜信号传递中的各种作用方面取得了进展,但很可能我们在识别它们的全部功能作用方面还只是触及了表面。
视网膜上的缝隙连接以及它们如何影响感受野的大小
脊椎动物的视网膜包含五个主要的神经元类:光感受器(视杆细胞和视锥细胞)、水平细胞、双极细胞、无腺细胞和神经节细胞。对于每一类神经元,我们描述了电和化学突触连接模式(图1).对于电突触,当已知时,我们概述了参与形成间隙连接的连接蛋白亚型,并描述了它们的生物物理特性。此外,我们尽可能地描述神经元对之间间隙连接的强度,用连接电导或耦合系数(即连接后细胞的膜电位变化除以连接前细胞的膜电位变化时,连接前细胞的膜电位变化)表示。
尽管我们将在下面概述耦合强度的特征,但重要的是要记住,通过间隙结的电导不是静态的,电导的变化可以对电突触在神经元电路中发挥的作用产生重大影响。例如,改变缝隙连接耦合强度可以通过改变视网膜神经元兴奋性感受野包围的强度和范围,显著改变视网膜神经元的感受野大小。对于视网膜上的许多神经元来说,光适应或昼夜节律可以调节电耦合的强度,从而改变感受野的大小。耦合强度的改变也可能改变视网膜回路中缝隙连接的其他作用。虽然耦合强度变化的分子机制已经被详细研究过(8),但在下一节中,我们将重点讨论耦合强度对感受野大小的影响。
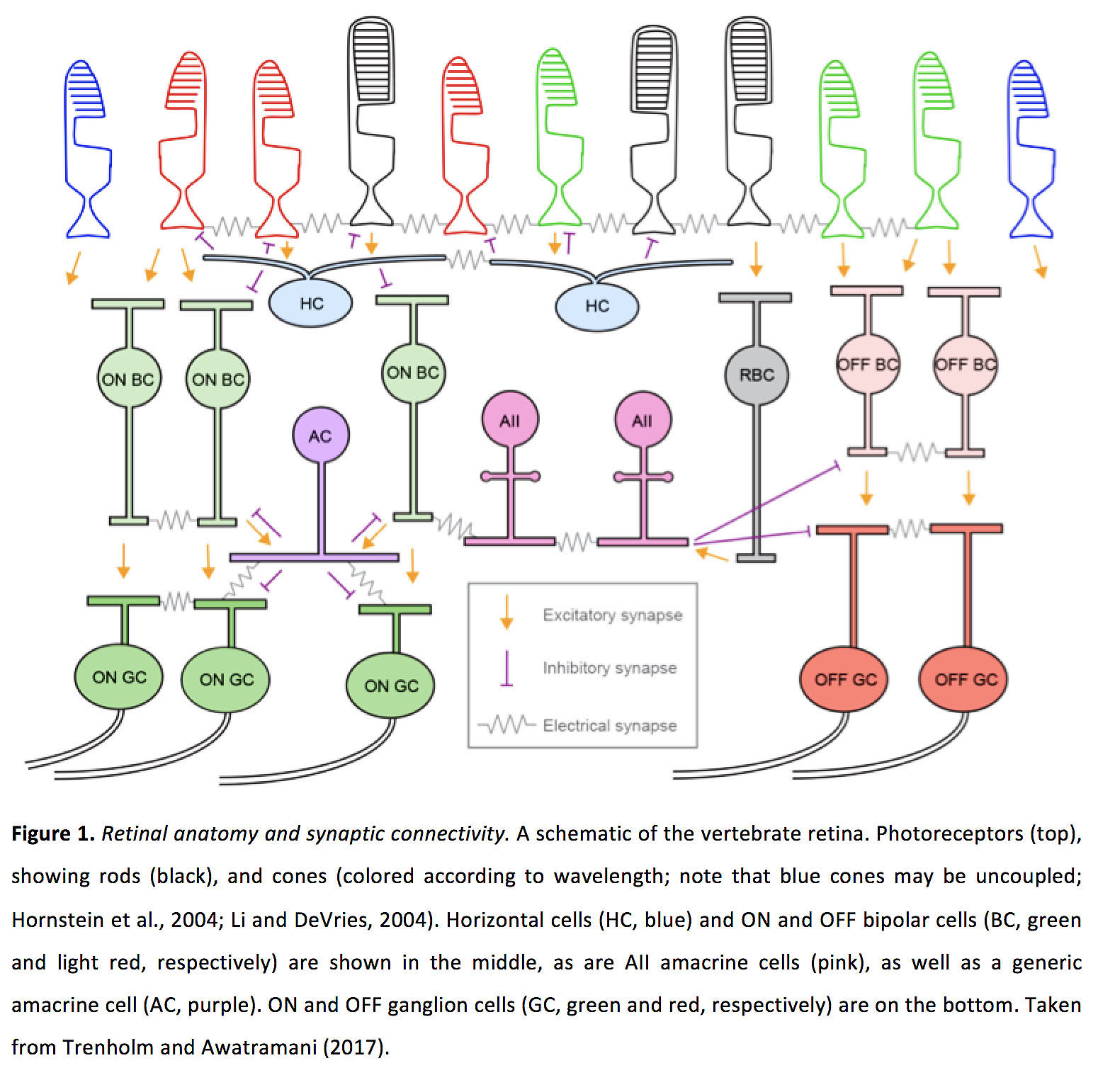
图1所示。视网膜解剖和突触连通性。脊椎动物视网膜的示意图光感受器(上)显示杆状物(黑色)和视锥细胞,根据波长而着色。注意,蓝色锥可能是解耦的(14,27)。中间是水平双极细胞(HC,蓝色)、ON双极细胞(ON BC,绿色)、OFF双极细胞(OFF BC,淡红色)和杆状双极细胞(RBC,灰色),以及全细胞型双极细胞(粉色)和普通双极细胞(AC,紫色)。ON和OFF神经节细胞(ON GC,绿色;OFF GC红色)在底部。摘自Trenholm and Awatramani(2017)(197)。
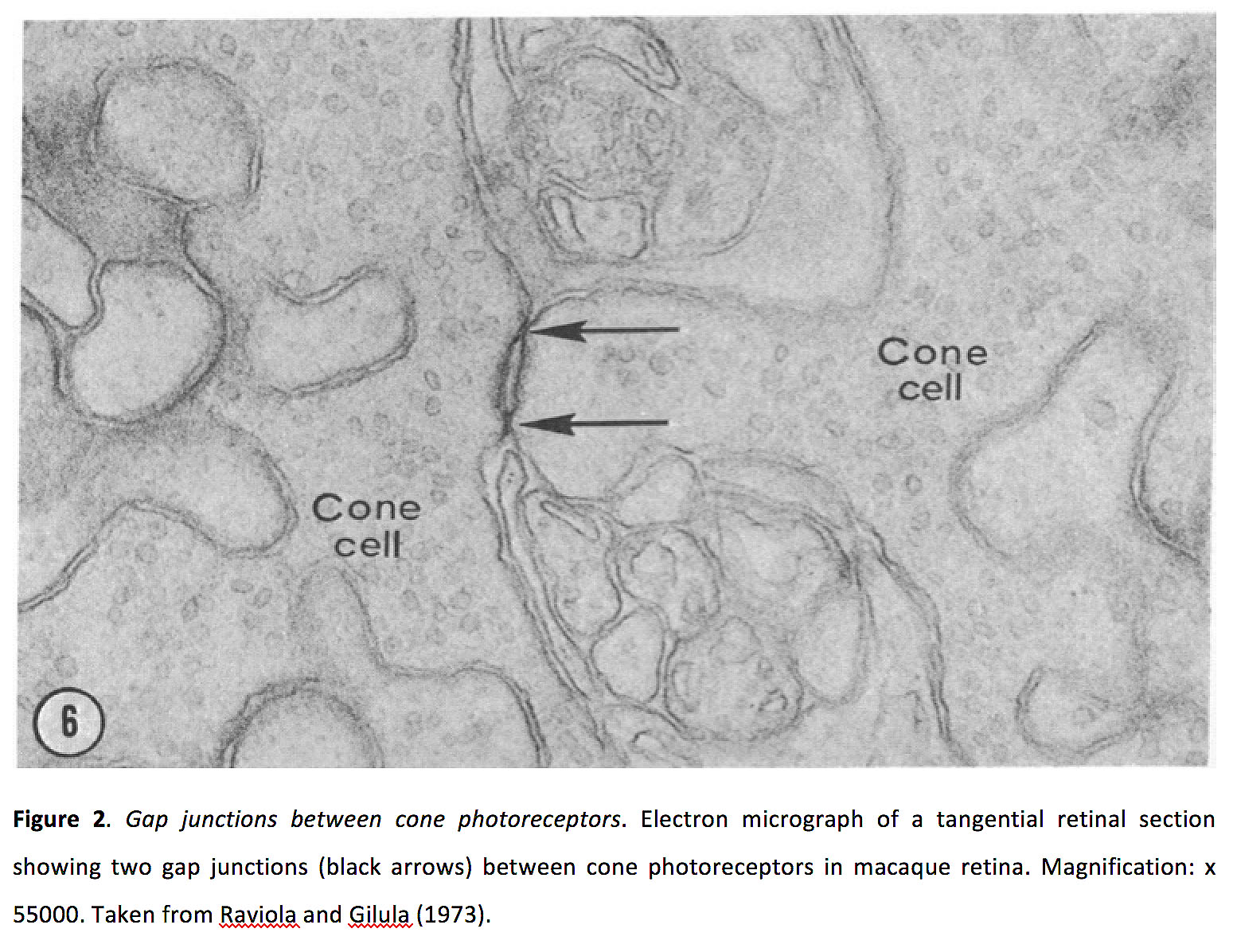 图2.锥体光感受器之间的缝隙连接.视网膜切线电子显微图显示猕猴视网膜锥状光感受器之间有两个间隙连接(黑色箭头)。放大:x 55000。摘自《拉维奥拉和吉鲁拉》(1973)(16)。
图2.锥体光感受器之间的缝隙连接.视网膜切线电子显微图显示猕猴视网膜锥状光感受器之间有两个间隙连接(黑色箭头)。放大:x 55000。摘自《拉维奥拉和吉鲁拉》(1973)(16)。
光感受器
视觉开始于杆状和锥状感光体,因为光子被转化为电化学信号(参见Webvision《杆和锥的光转导》章节).光感受器与水平和双极细胞产生谷氨酸突触(保存离子信号的突触与水平细胞和OFF双极细胞;与ON双极细胞的代谢唱歌倒置突触)。光感受器也形成同源的(即锥-锥和杆-杆)和异源的(杆-锥)电突触(图2)(9-17)。
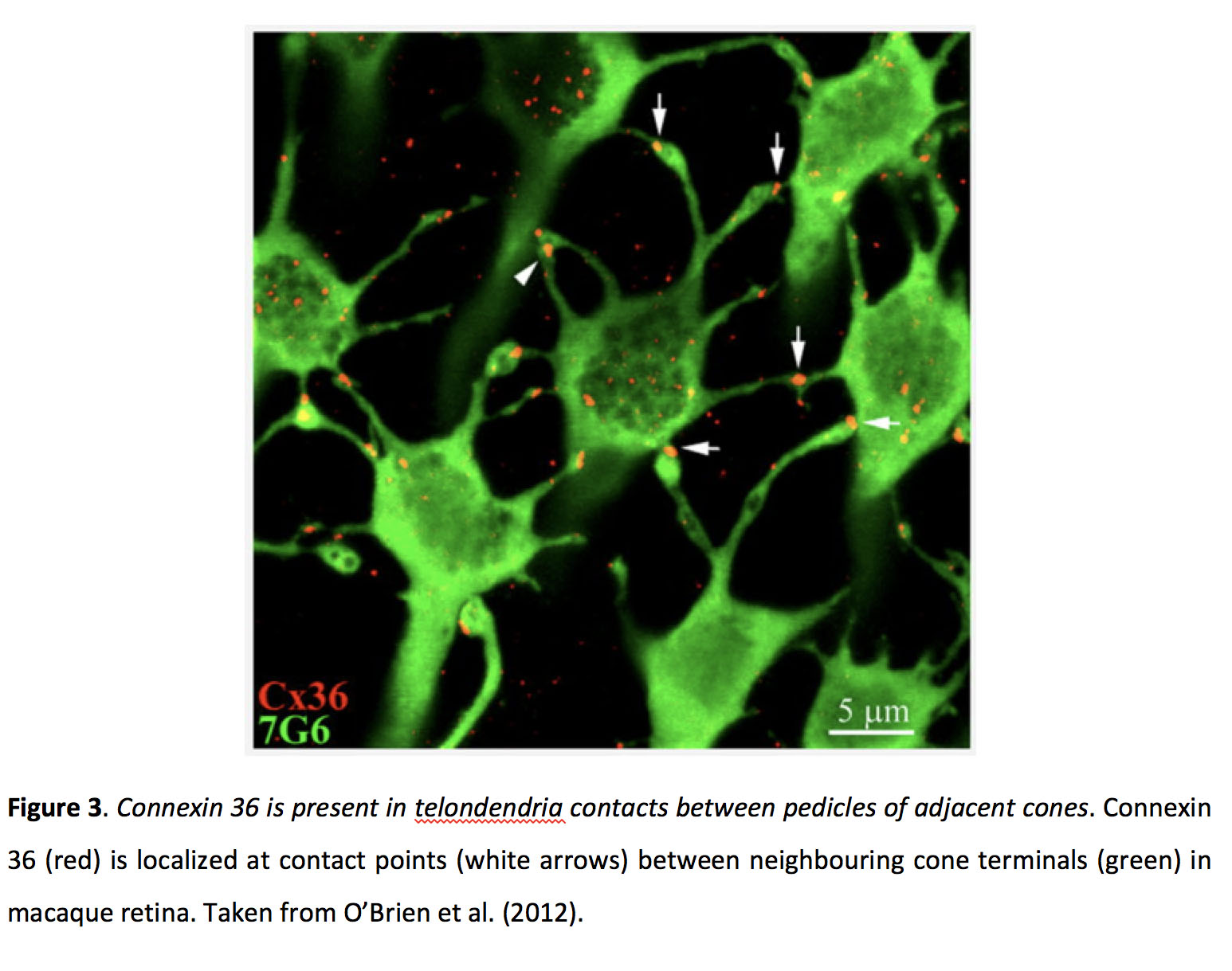 图3.连接蛋白36存在于相邻锥椎弓根之间的近突接触中.连接蛋白36 (Cx36,红色)位于猕猴视网膜相邻锥体末端(7G6,绿色,锥体抑制素抗体)之间的接触点(白色箭头)。摘自O 'Brien等人(2012)(21)。
图3.连接蛋白36存在于相邻锥椎弓根之间的近突接触中.连接蛋白36 (Cx36,红色)位于猕猴视网膜相邻锥体末端(7G6,绿色,锥体抑制素抗体)之间的接触点(白色箭头)。摘自O 'Brien等人(2012)(21)。
哺乳动物锥体间隙连接含有连接蛋白36 (Cx36) (图3)(18-21)和Cx35在斑马鱼(22),而涉及杆状电突触的连接蛋白尚未被明确识别(22-24),杆状和锥状突触之间的耦合似乎依赖于Cx36(25)。光感受器间隙连接位于视锥细胞轴突末端周围(11,16,26 -28)。
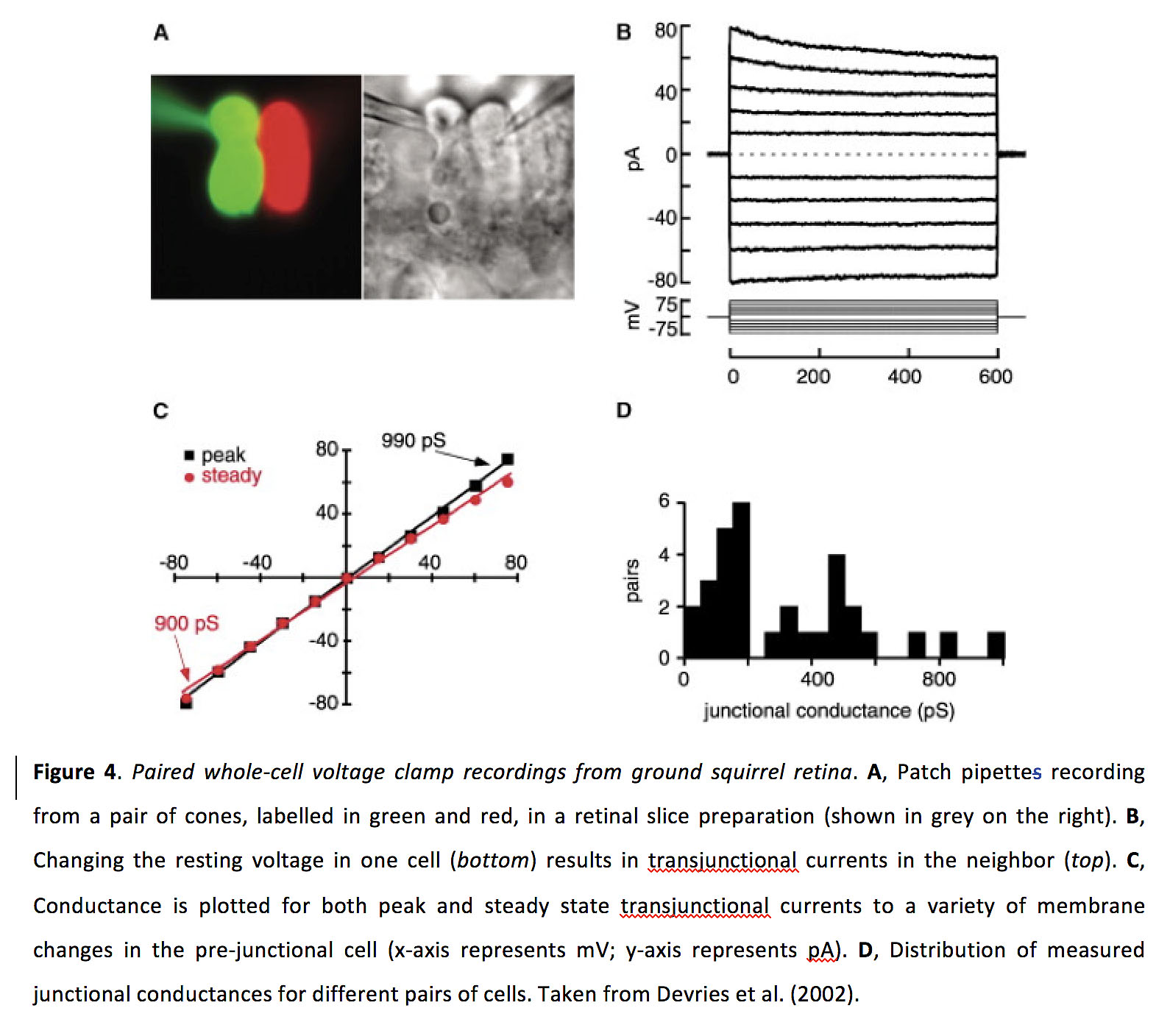 图4.来自地鼠视网膜的配对全细胞电压钳记录.一个,贴片移液管从一对视锥细胞(标记为绿色和红色)记录视网膜切片制剂(显示为右侧灰色)。B,改变单个电池的静息电压(底)导致相邻(前).C,分别绘制跨接电流峰值和稳态到预连接细胞内各种膜变化时的电导(x轴表示mV;轴代表pA)。D,不同电池对的实测结电导分布。摘自Devries等人(2002)(12)。
图4.来自地鼠视网膜的配对全细胞电压钳记录.一个,贴片移液管从一对视锥细胞(标记为绿色和红色)记录视网膜切片制剂(显示为右侧灰色)。B,改变单个电池的静息电压(底)导致相邻(前).C,分别绘制跨接电流峰值和稳态到预连接细胞内各种膜变化时的电导(x轴表示mV;轴代表pA)。D,不同电池对的实测结电导分布。摘自Devries等人(2002)(12)。
根据生理记录,同源光感受器耦合似乎比异源耦合更强。在200-800 pS (图4)(12,14,15,27,29),耦合系数约为0.15(14,30)。据报道,杆锥间隙结的结电导是双向对称的,在40-200 pS(31)范围内,耦合系数约为0.04(32)。从将染料注入视锥细胞的示踪染料实验来看,与视杆细胞偶联比只与视锥细胞偶联或与视杆细胞和视锥细胞同时偶联更常见,有些视锥细胞可能是不偶联的(13)。相比之下,在示踪染料实验中,染料注入到棒中,棒-棒联轴器似乎比棒-锥联轴器更常见,一些棒可能是解耦的(15)。
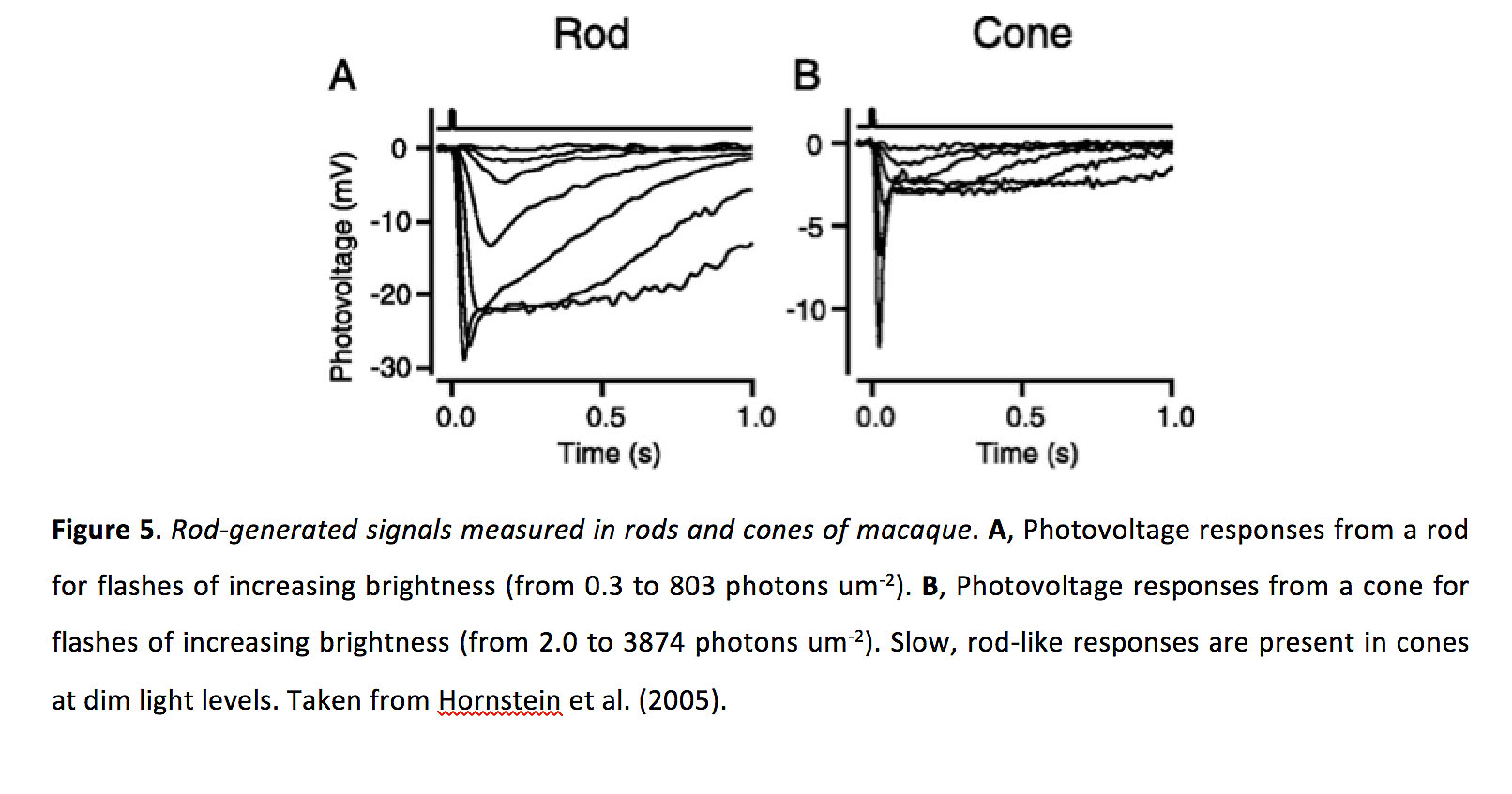 图5.猕猴的视杆细胞和视锥细胞产生的信号.一个,光棒对亮度增加的闪光(从0.3到803光子um)的光电压响应-2).B,锥体对亮度增加(从2.0到3874光子)的光电压响应-2).在昏暗的光线下,视锥细胞会出现缓慢的杆状反应。激发波长为500 nm。摘自Hornstein等人(2005)(13)。
图5.猕猴的视杆细胞和视锥细胞产生的信号.一个,光棒对亮度增加的闪光(从0.3到803光子um)的光电压响应-2).B,锥体对亮度增加(从2.0到3874光子)的光电压响应-2).在昏暗的光线下,视锥细胞会出现缓慢的杆状反应。激发波长为500 nm。摘自Hornstein等人(2005)(13)。
昼夜节律似乎是改变鱼和老鼠视网膜中光感受器电耦合强度的主要控制器,而不是光适应(33)。在人类身上进行的心理物理实验表明,锥体耦合似乎不受光线适应的影响,但昼夜节律的影响尚未得到测试(12)。相反,光适应,而不是昼夜节律,似乎调节蝾螈光感受器之间的电耦合强度(31)。改变缝隙连接的强度可以改变光感受器的感受野大小。例如,在视杆和视锥的电耦合更强的状态下,视锥有更大的接收场(33)。类似地,当它们耦合得更强时,棒状接受场也会更大,而耦合降低了它们对昏暗闪光响应的反复试验的变异性(15)。最后,异型间隙连接允许锥响应传递到杆(34),杆响应传递到锥(34)。图5)(13,35 -37),这意味着电耦合的增加应该会扩大给定杆或锥所能响应的光强度范围,至少对锥来说,这种耦合可以导致阈值响应的可靠性增加。
水平细胞
横向细胞从光感受器接收谷氨酸输入,然后向光感受器提供负反馈,并可能对双极细胞产生前馈抑制(参见Webvision章节s -potential和Horizontal cell).反馈相互作用的机制仍在积极研究中(38)。此外,水平细胞与缝隙连接紧密相连(图639)(26日)。
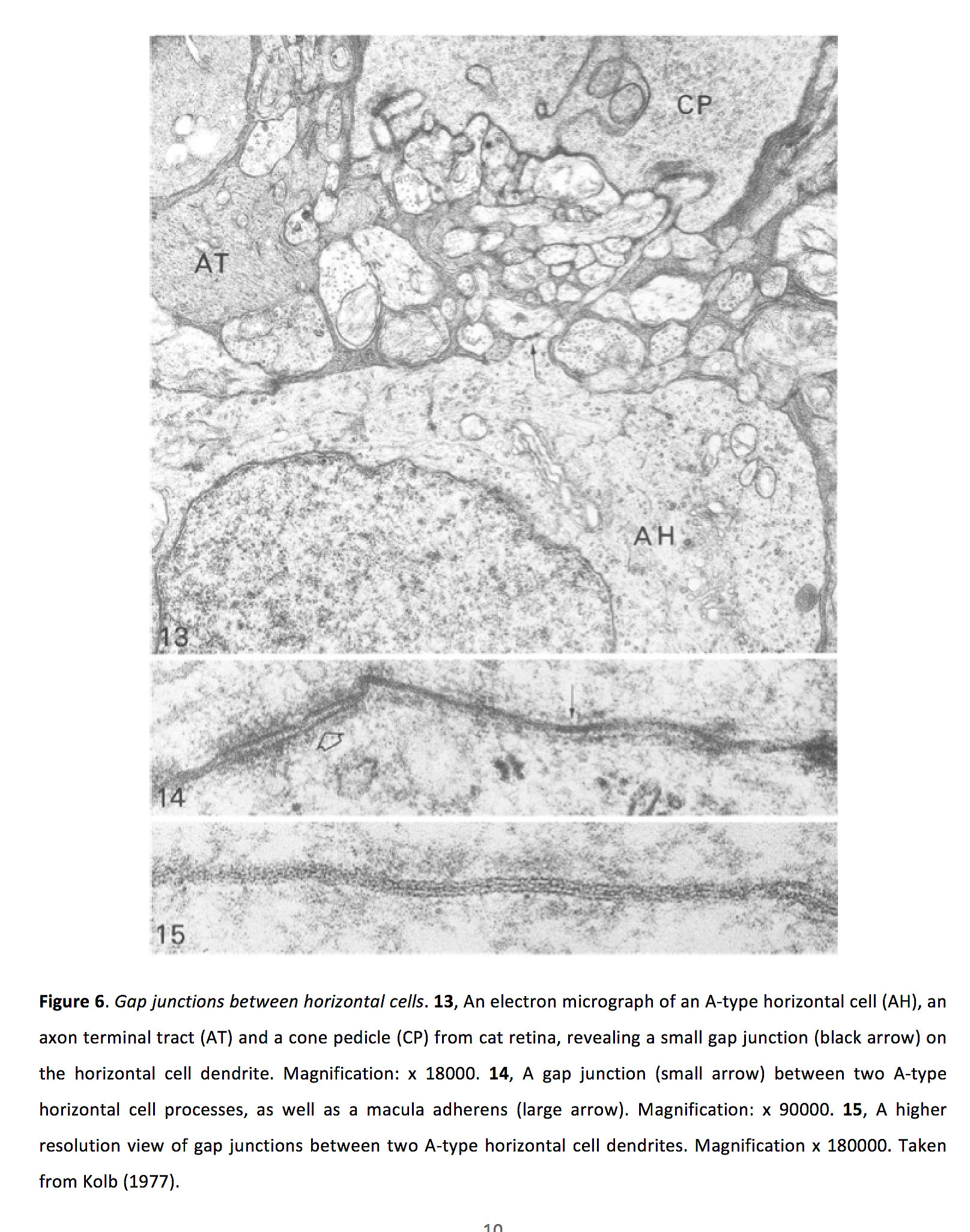 图6.水平细胞之间的间隙连接.13猫视网膜上的a型水平细胞(AH)、轴突末端束(AT)和锥体蒂(CP)的电子显微镜照片,显示水平细胞树突上有一个小间隙连接(黑色箭头)。放大:x 18000。14两个A型水平细胞突起之间的间隙连接(小箭头),以及黄斑粘附(大箭头)。放大:x 90000。15,两个A型水平细胞树突之间的间隙连接的更高分辨率视图。放大x 180000。摘自Kolb(1977)(26)。
图6.水平细胞之间的间隙连接.13猫视网膜上的a型水平细胞(AH)、轴突末端束(AT)和锥体蒂(CP)的电子显微镜照片,显示水平细胞树突上有一个小间隙连接(黑色箭头)。放大:x 18000。14两个A型水平细胞突起之间的间隙连接(小箭头),以及黄斑粘附(大箭头)。放大:x 90000。15,两个A型水平细胞树突之间的间隙连接的更高分辨率视图。放大x 180000。摘自Kolb(1977)(26)。
水平细胞间隙连接既可以是轴突的,也可以是树突的。在小鼠中,Cx57存在于两种间隙连接中(图7)(40,41),而Cx50定位于轴突间隙连接(42)。在兔a型水平细胞中,Cx50定位于树突间隙连接(43),而在b型水平细胞中,Cx57定位于轴突间隙连接(44)。在斑马鱼中,水平细胞间隙连接似乎由Cx52.6, 52.9和55.5组成(45,46)。
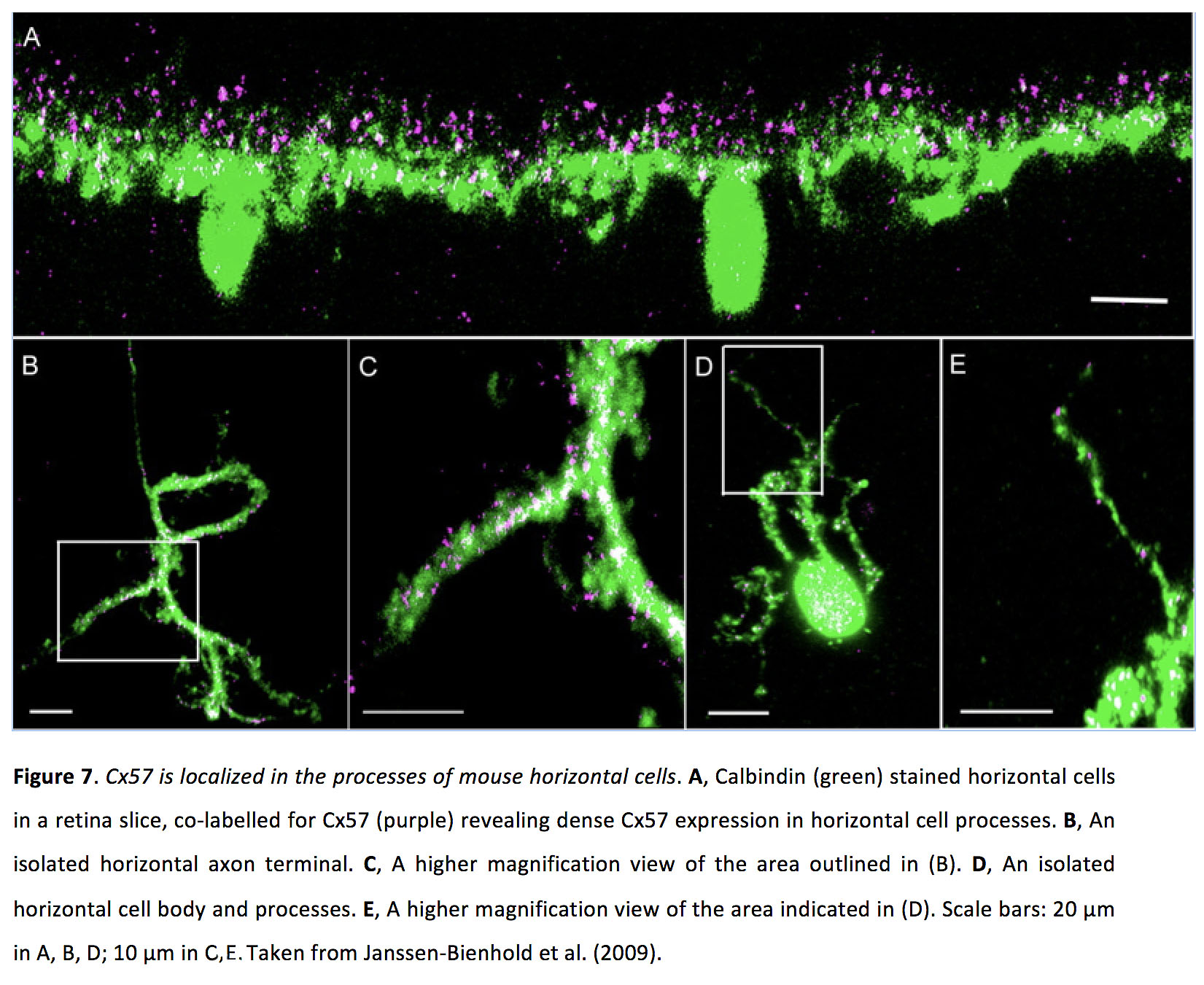 图7.Cx57定位于小鼠水平细胞过程中.一个在视网膜切片上,Calbindin(绿色)染色的水平细胞,共同标记Cx57(紫色)显示水平细胞过程中密集的Cx57表达。B孤立的水平轴突末端。C, (B)中所述区域的放大图。D孤立的水平细胞体和细胞突起。E, (D)所示区域的高倍放大视图。比例尺:A、B、D的20µm;C, e中10 μ m取自Janssen-Bienhold等人(2009)(41)。
图7.Cx57定位于小鼠水平细胞过程中.一个在视网膜切片上,Calbindin(绿色)染色的水平细胞,共同标记Cx57(紫色)显示水平细胞过程中密集的Cx57表达。B孤立的水平轴突末端。C, (B)中所述区域的放大图。D孤立的水平细胞体和细胞突起。E, (D)所示区域的高倍放大视图。比例尺:A、B、D的20µm;C, e中10 μ m取自Janssen-Bienhold等人(2009)(41)。
水平细胞之间的耦合似乎非常强,双向对称的连接电导为> 1ns,在培养的水平细胞中耦合系数为0.5或更大(47-50)。从引人注目的示踪染料实验中,将染料注入到单个水平细胞中,发现染料可以扩散到周围1000多个水平细胞中,延伸的距离超过1毫米(图8)(51)。
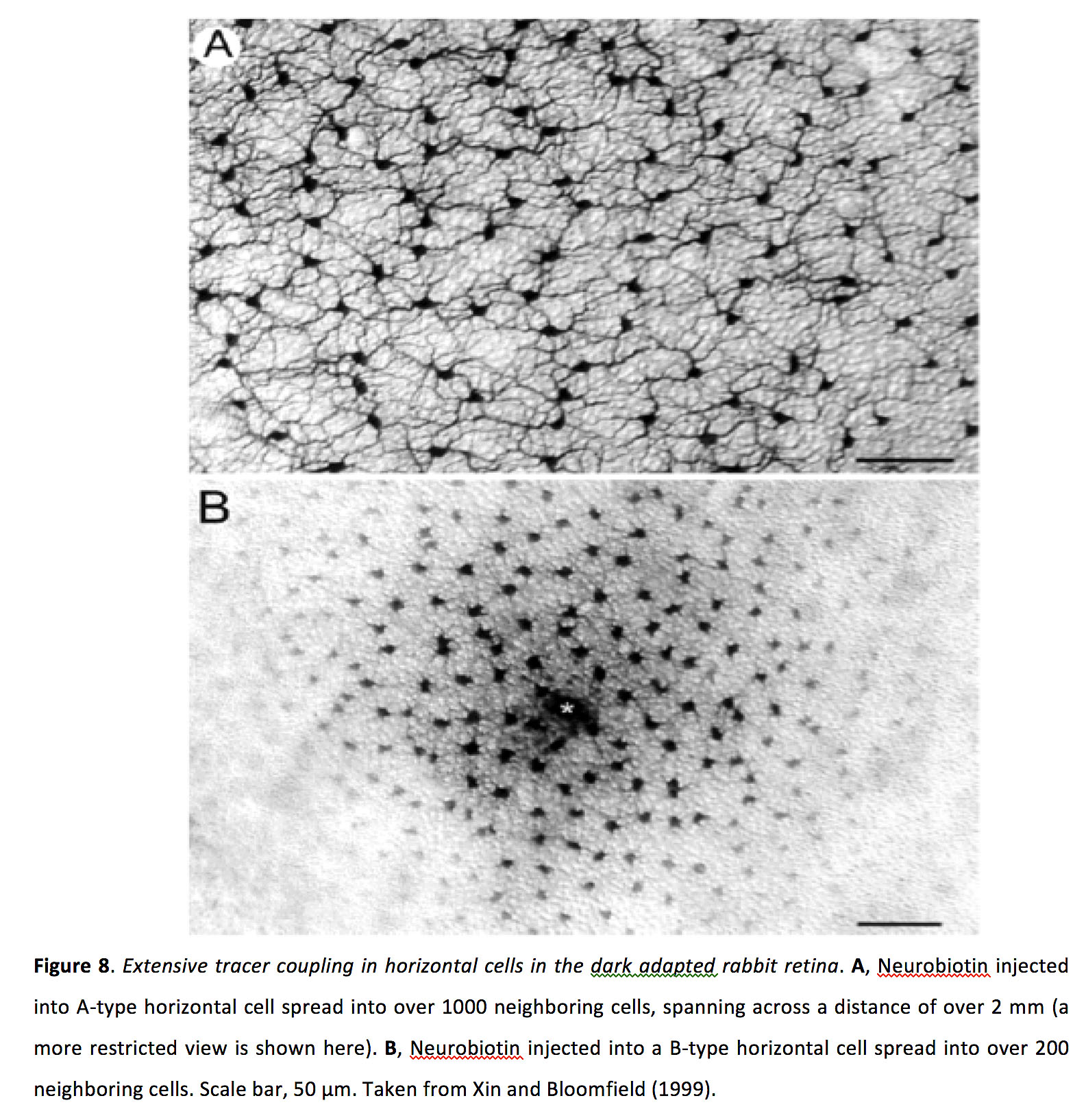 图8.在适应黑暗的兔子视网膜水平细胞中广泛的示踪剂偶联.一个注射到a型水平细胞的神经生物素扩散到超过1000个相邻的细胞中,跨越超过2毫米的距离(这里显示的是一个更有限的视图)。B注入b型水平细胞的神经生物素扩散到200多个邻近细胞。比例尺,50µm。摘自Xin和Bloomfield(1999)(51)。
图8.在适应黑暗的兔子视网膜水平细胞中广泛的示踪剂偶联.一个注射到a型水平细胞的神经生物素扩散到超过1000个相邻的细胞中,跨越超过2毫米的距离(这里显示的是一个更有限的视图)。B注入b型水平细胞的神经生物素扩散到200多个邻近细胞。比例尺,50µm。摘自Xin和Bloomfield(1999)(51)。
在水平电池中,光适应似乎是控制缝隙连接耦合强度的主要因素(52,53)。水平细胞耦合已被证明显示出一种依赖于亮度水平的三相关系:示踪剂耦合实验表明,在暗适应或“暗抑制”状态下耦合最小;在所谓的“光敏”状态中,在中等光水平下,示踪剂耦合大大增加;在光适应状态下,示踪剂耦合非常弱(图9)(51)。
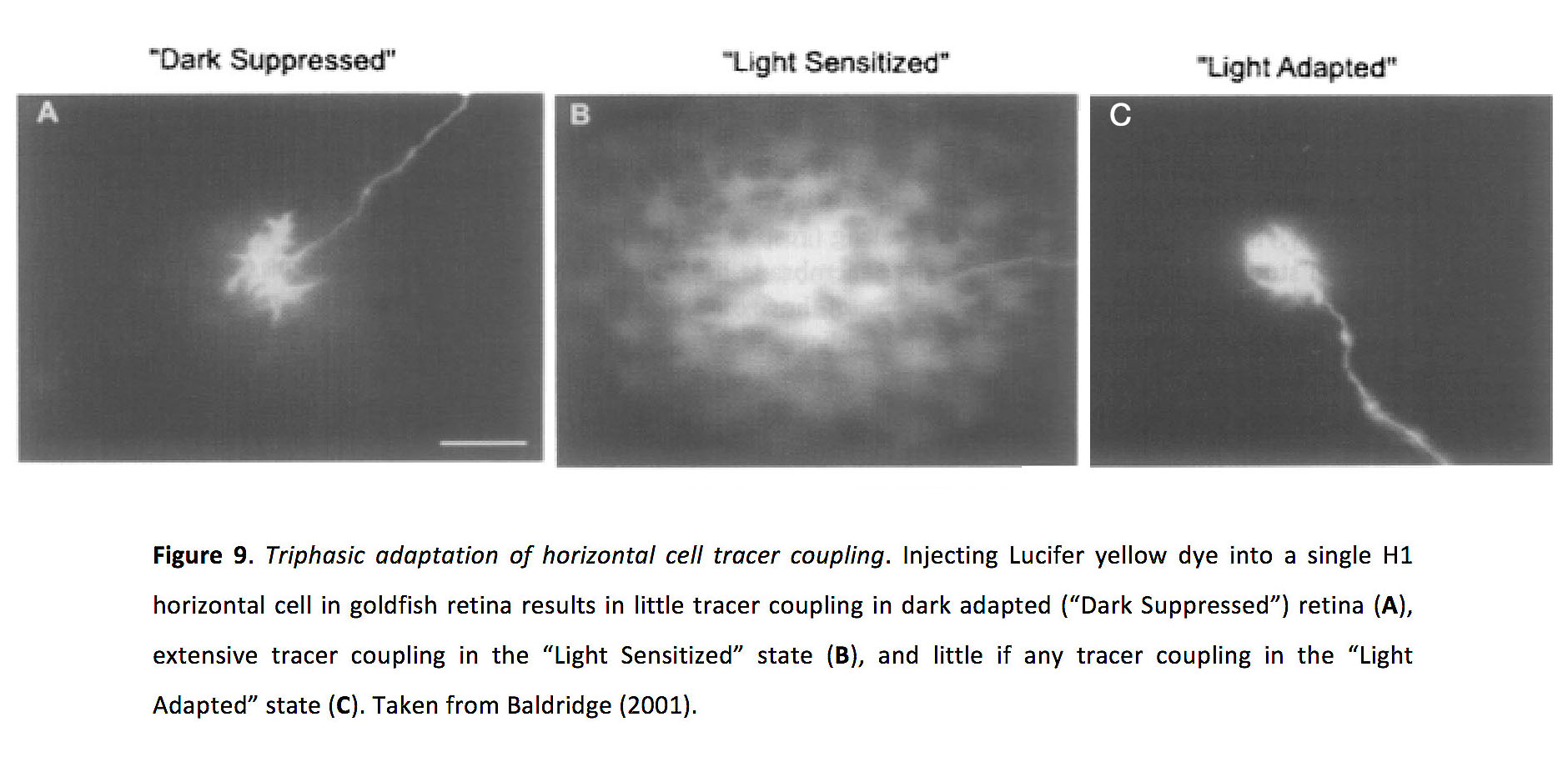 图9.水平细胞示踪剂耦合的三相适应.将路西法黄染料注射到金鱼视网膜的单个H1水平细胞中,会导致暗适应(“暗抑制”)视网膜中的微量示踪剂偶联(一个),“光致敏”状态下的大量示踪剂耦合(B),并且在“光适应”状态下几乎没有任何跟踪耦合(C).摘自Baldridge(2001)(198)。
图9.水平细胞示踪剂耦合的三相适应.将路西法黄染料注射到金鱼视网膜的单个H1水平细胞中,会导致暗适应(“暗抑制”)视网膜中的微量示踪剂偶联(一个),“光致敏”状态下的大量示踪剂耦合(B),并且在“光适应”状态下几乎没有任何跟踪耦合(C).摘自Baldridge(2001)(198)。
水平细胞耦合似乎会强烈影响感受野的大小(54),在光敏状态下,感受野大约是光敏状态的三倍(51)。与这一发现一致的是,敲除小鼠Cx57会减少水平细胞感受野的大小(图10)(55)。
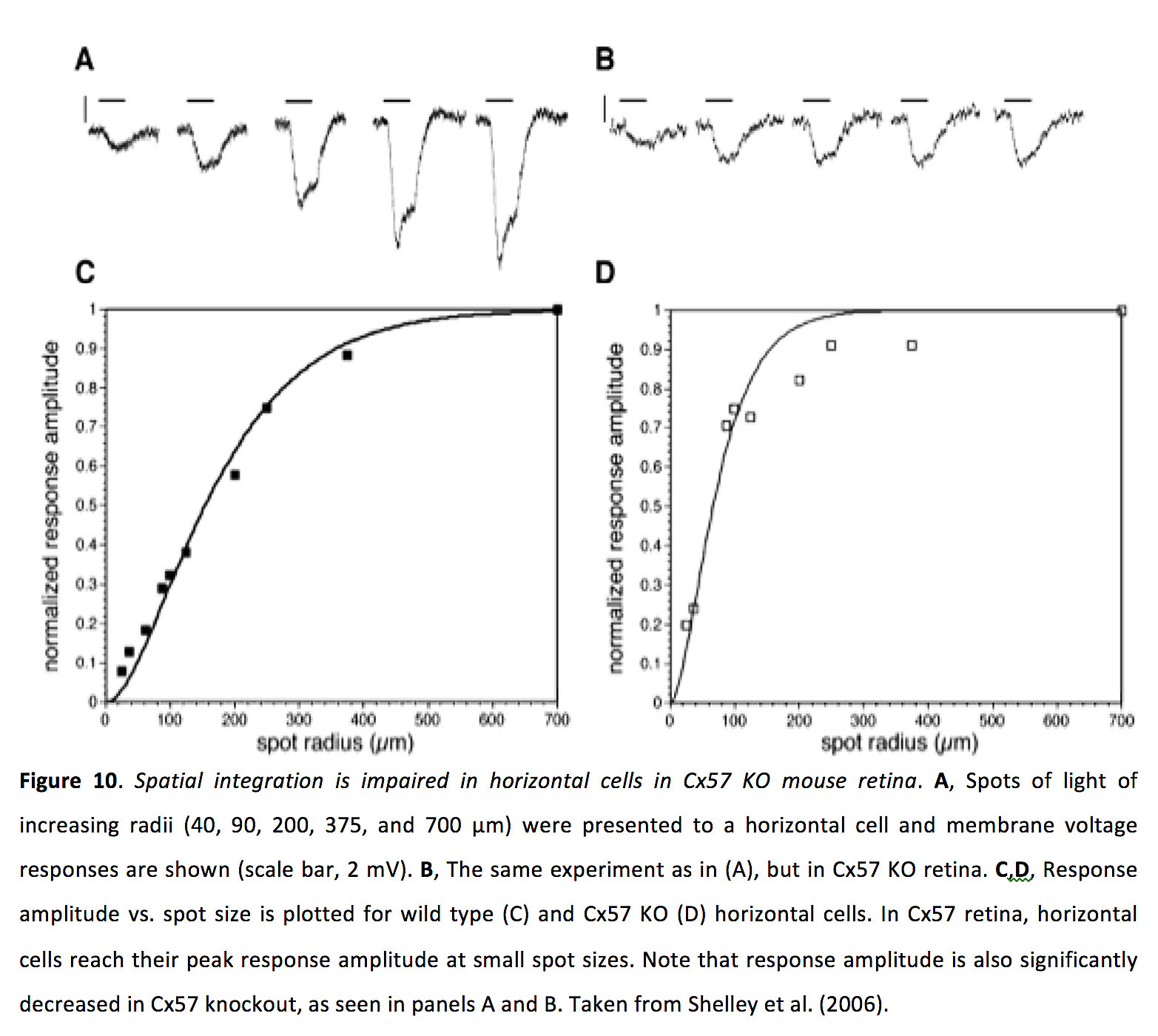 图10.cx57ko小鼠视网膜水平细胞空间整合受损.一个在水平细胞上出现了半径增加的光斑(40、90、200、375和700 μ m),并显示了膜电压响应(比较尺,2 mV)。B,与(A)相同的实验,但Cx57 KO视网膜。C,D,对野生型(C)和Cx57 KO (D)水平细胞绘制了响应振幅与光斑大小的关系。在Cx57视网膜中,水平细胞在小斑点尺寸时达到峰值响应振幅。请注意,敲除Cx57基因后的响应振幅也显著降低,如图A和b所示。摘自Shelley等人(2006)(55)。
图10.cx57ko小鼠视网膜水平细胞空间整合受损.一个在水平细胞上出现了半径增加的光斑(40、90、200、375和700 μ m),并显示了膜电压响应(比较尺,2 mV)。B,与(A)相同的实验,但Cx57 KO视网膜。C,D,对野生型(C)和Cx57 KO (D)水平细胞绘制了响应振幅与光斑大小的关系。在Cx57视网膜中,水平细胞在小斑点尺寸时达到峰值响应振幅。请注意,敲除Cx57基因后的响应振幅也显著降低,如图A和b所示。摘自Shelley等人(2006)(55)。
由于水平细胞向光感受器提供负反馈,有人认为它们可能负责在下游神经节细胞中产生抑制性周围感受野(56,57),在这种情况下,修改水平细胞耦合强度将修改神经节细胞感受野大小。然而,水平细胞反馈对神经节细胞周围反应的影响仍不确定,至少在某些情况下,水平细胞不驱动神经节细胞明显的周围反应(58-60)。此外,尽管降低了水平细胞感受野的大小,敲除Cx57对小鼠神经节细胞的空间调节和视力几乎没有影响(61)。相反,水平细胞可能在神经节细胞的时间调节中起作用。例如,有人认为强电耦合可能通过有效分流来自光感受器的输入电流,从功能上移除视网膜电路中的水平细胞。在这种情况下,改变耦合强度可以使视网膜从日光条件下的短整合时间模式切换到夜光条件下的长整合时间模式(62)。在另一项研究中,水平细胞对光感受器的反馈被证明对神经节细胞反应的短暂和/或持续部分有不同的影响(58),这可能提供了另一种机制,即水平细胞耦合的变化可能影响神经节细胞的时间调谐。
双极细胞
双极细胞,其中至少有10种(63种),将光感受器信号传递到谷氨酸突触的无分泌细胞和神经节细胞(参见Webvision章节脊椎动物视网膜中的双极细胞通路).此外,双极细胞可与其他双极细胞和无分泌细胞电偶联。最常见的是,双极细胞偶联到所有的无分泌细胞(图11)(64),尽管最近的研究表明某些ON锥双极细胞与非所有的无分泌细胞形成间隙连接(59,65)。参见Webvision关于所有无分泌细胞的章节.
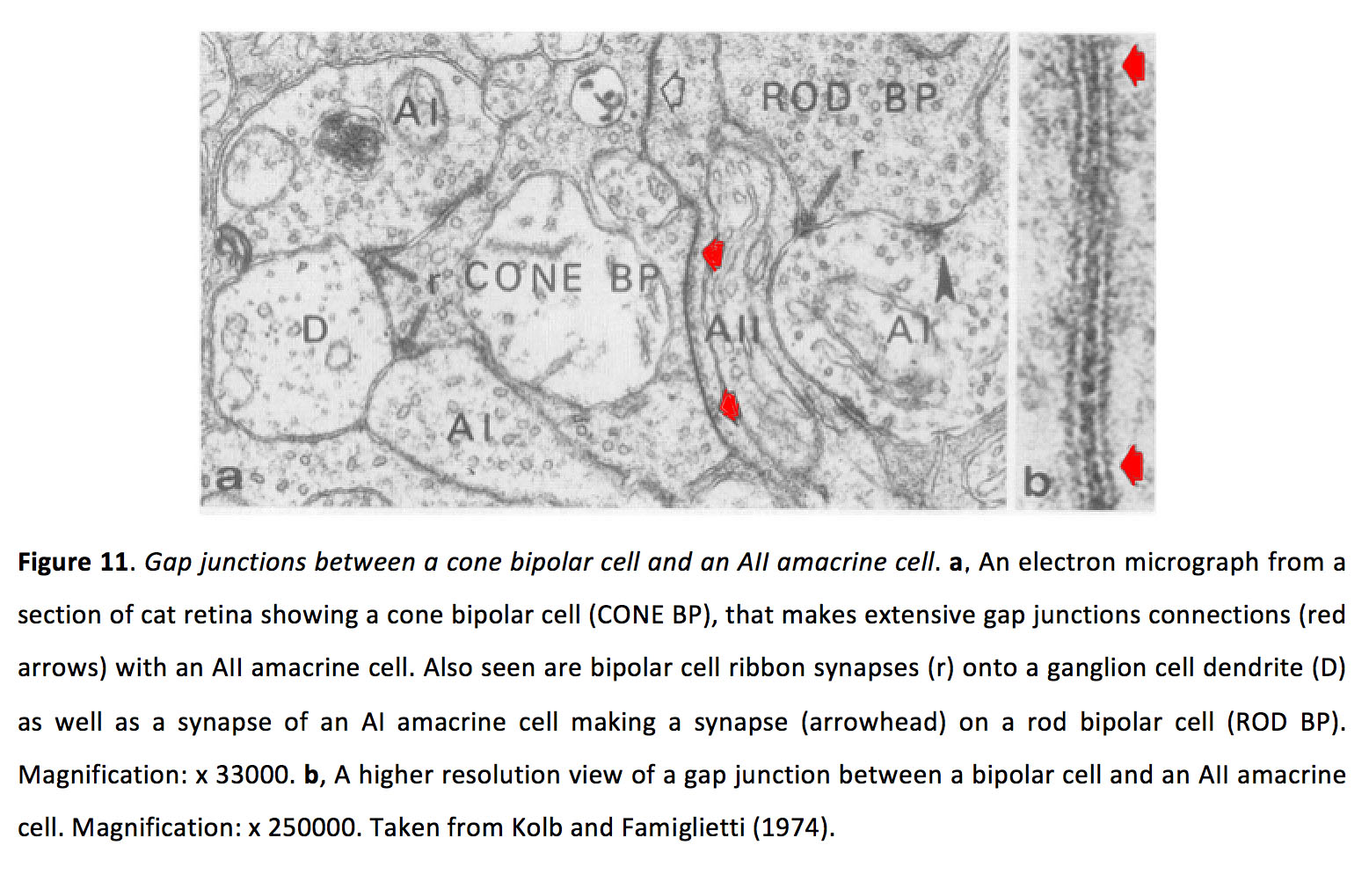 图11.锥体双极细胞与全无分泌细胞之间的间隙连接。一个猫视网膜切片的电子显微照片显示一个锥双极细胞(cone BP),它与一个全aiamacrine细胞形成广泛的间隙连接(红色箭头)。还可以看到双极细胞带突触(r)在神经节细胞树突(D)上,以及AI无分泌细胞的突触(箭头)在杆状双极细胞(rod BP)上形成突触(箭头)。人工智能也被称为A17。放大:x 33000。b双极细胞和全无分泌细胞之间的间隙连接的高分辨率视图。放大:x 250000。摘自Kolb和Famiglietti(1974)(64)。
图11.锥体双极细胞与全无分泌细胞之间的间隙连接。一个猫视网膜切片的电子显微照片显示一个锥双极细胞(cone BP),它与一个全aiamacrine细胞形成广泛的间隙连接(红色箭头)。还可以看到双极细胞带突触(r)在神经节细胞树突(D)上,以及AI无分泌细胞的突触(箭头)在杆状双极细胞(rod BP)上形成突触(箭头)。人工智能也被称为A17。放大:x 33000。b双极细胞和全无分泌细胞之间的间隙连接的高分辨率视图。放大:x 250000。摘自Kolb和Famiglietti(1974)(64)。
双极细胞之间的间隙连接可以定位在树突(66)或轴突末端(67,68),而ON锥双极细胞和所有无分泌细胞之间的间隙连接定位在双极细胞轴突末端(69)。在小鼠视网膜中,双极细胞间隙连接包含Cx36或Cx45 (图12)(70 - 73)。
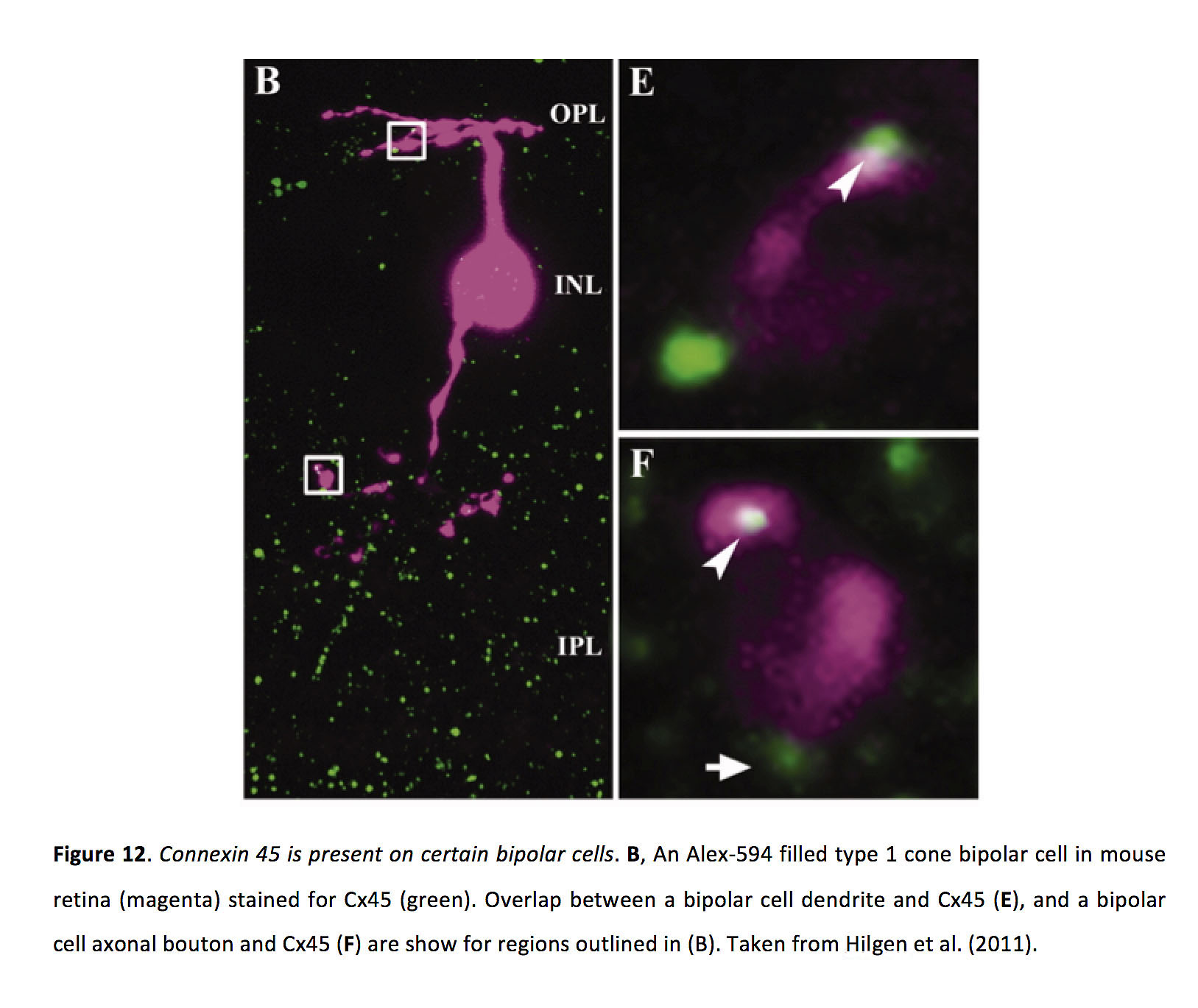 图12.连接蛋白45存在于某些双极细胞中.B小鼠视网膜中Alexa-594填充型1型锥体双极细胞(洋红色)为Cx45(绿色)染色。双极细胞树突与Cx45的重叠(E)和双极细胞轴突钮扣和Cx45 (F)显示了(B)中概述的区域。摘自Hilgen等人(2011)(72)。
图12.连接蛋白45存在于某些双极细胞中.B小鼠视网膜中Alexa-594填充型1型锥体双极细胞(洋红色)为Cx45(绿色)染色。双极细胞树突与Cx45的重叠(E)和双极细胞轴突钮扣和Cx45 (F)显示了(B)中概述的区域。摘自Hilgen等人(2011)(72)。
双极细胞-双极细胞间隙连接是双向的、对称的,其连接电导约为1ns,耦合系数约为0.3 (图13)(66年,74 - 77)。
锥体双极细胞与全锥体无分泌细胞形成双向但不对称的缝隙连接,尽管这种测量到的不对称可能是输入电阻差异的假象。ON锥双极细胞和AII无轴细胞之间的结电导约为1ns,耦合系数在0.3-0.6之间(图14)(64,70,78 -80)。
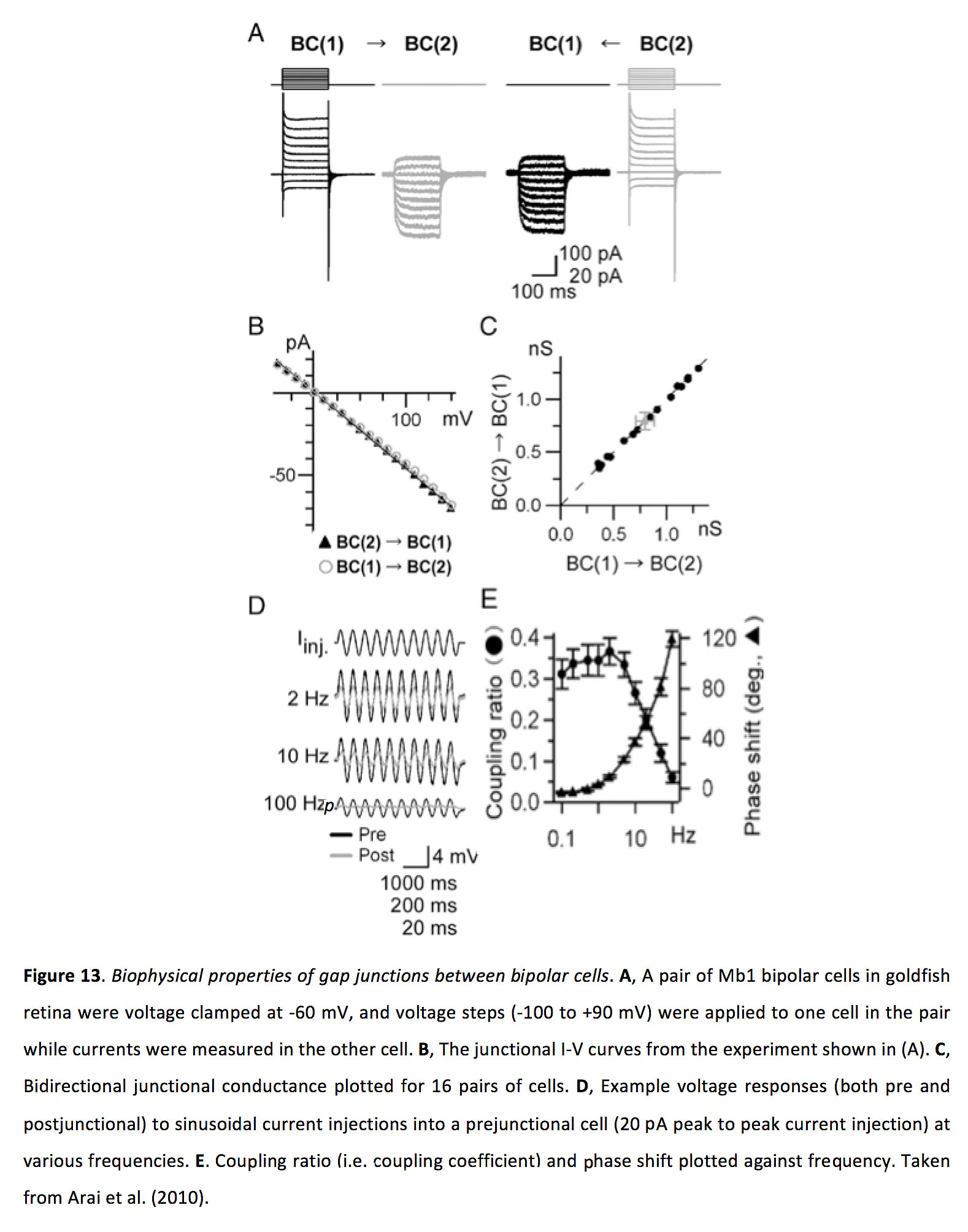 图13.双极细胞间隙连接的生物物理特性.一个对金鱼视网膜上的一对Mb1双极细胞在-60 mV电压下钳住,对其中一个细胞施加电压步进(-100到+90 mV),同时在另一个细胞中测量电流。B, (A)所示实验的结I-V曲线。C, 16对细胞的双向结电导图。D例电压响应(包括连接前和连接后)正弦电流注入到连接前电池(20pa峰值对峰值电流注入)在不同频率。E,耦合比(即耦合系数)和相移与频率的关系。摘自Arai等人(2010)(66)。
图13.双极细胞间隙连接的生物物理特性.一个对金鱼视网膜上的一对Mb1双极细胞在-60 mV电压下钳住,对其中一个细胞施加电压步进(-100到+90 mV),同时在另一个细胞中测量电流。B, (A)所示实验的结I-V曲线。C, 16对细胞的双向结电导图。D例电压响应(包括连接前和连接后)正弦电流注入到连接前电池(20pa峰值对峰值电流注入)在不同频率。E,耦合比(即耦合系数)和相移与频率的关系。摘自Arai等人(2010)(66)。
在光适应状态下,电连接的双极细胞之间的间隙结电导增加,至少如金鱼中Mb-1双极细胞所示(66)。虽然有人认为双极细胞耦合可以导致感受野大小的增加,一些研究发现其显著大于树突状场的大小(74,75,81),但也见(82),据我们所知,改变光适应状态对双极细胞耦合和感受野大小的影响从未被直接测试过。此外,由于先前的研究表明,光适应可以在双极细胞中产生拮抗的周围感受野(83),这种周围抑制可能会抵消邻近细胞通过缝隙连接输入提供的兴奋性周围反应,从而限制缝隙连接耦合扩大感受野大小的能力。因此,虽然双极细胞间隙连接似乎增强了它们整合空间分离刺激的能力,并优先增强了它们对低对比度刺激的信号传导(84),但还需要进一步研究来确定光适应对双极细胞间隙连接耦合强度的影响,以及检验这种变化是否有效地改变了感受场大小。
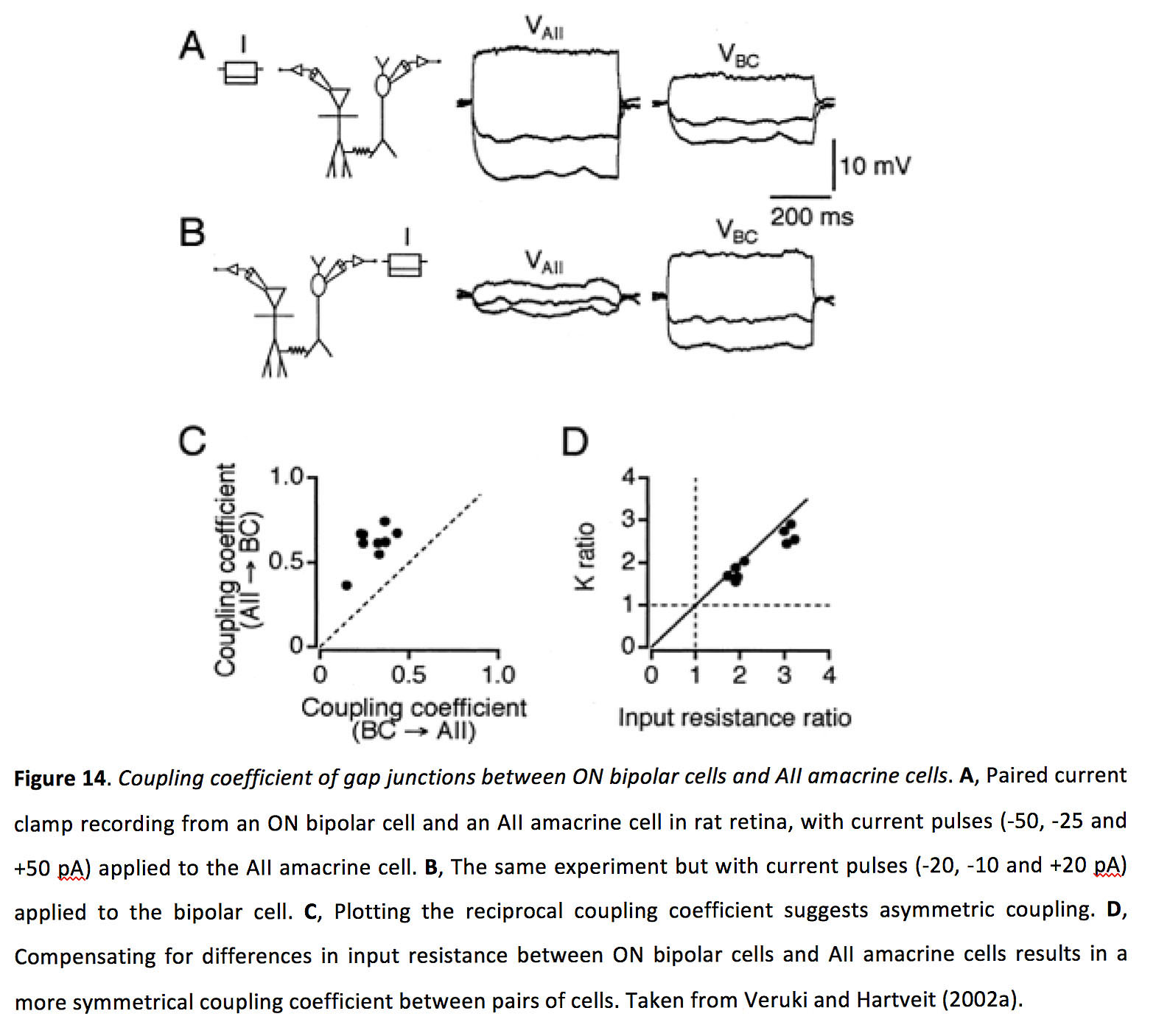 图14.ON极性细胞与A极性细胞间隙连接的耦合系数二世无长突细胞.一个配对电流钳记录大鼠视网膜上的ON双极细胞和all - amacrine细胞,对all - amacrine细胞施加电流脉冲(-50,-25和+50 pA)。B,相同的实验,但在双极电池上施加电流脉冲(-20,-10和+20 pA)。C,绘制倒数耦合系数表示非对称耦合。D,补偿ON双极细胞和aiamacrine细胞之间输入电阻的差异,导致对细胞之间更对称的耦合系数。取自Veruki和Hartveit (2002a)(199)。
图14.ON极性细胞与A极性细胞间隙连接的耦合系数二世无长突细胞.一个配对电流钳记录大鼠视网膜上的ON双极细胞和all - amacrine细胞,对all - amacrine细胞施加电流脉冲(-50,-25和+50 pA)。B,相同的实验,但在双极电池上施加电流脉冲(-20,-10和+20 pA)。C,绘制倒数耦合系数表示非对称耦合。D,补偿ON双极细胞和aiamacrine细胞之间输入电阻的差异,导致对细胞之间更对称的耦合系数。取自Veruki和Hartveit (2002a)(199)。
无长突细胞
Amacrine细胞,其中有超过45种类型(5,85),通过GABA或甘氨酸释放对双极细胞、其他Amacrine细胞和神经节细胞提供抑制。一些无分泌细胞也可以共同释放乙酰胆碱(86-88)、多巴胺(89)或谷氨酸(90)。许多无突细胞形成同源间隙连接(78,79,91 -93),以及与双极细胞(如上所述)和神经节细胞(94,95)形成异源间隙连接。所有无分泌细胞在其树突树枝中形成间隙连接(图15)(69)。似乎其他类型的无突细胞也与无突细胞和神经节细胞形成树突间隙连接,因为这通常是它们突起重叠的唯一位置。
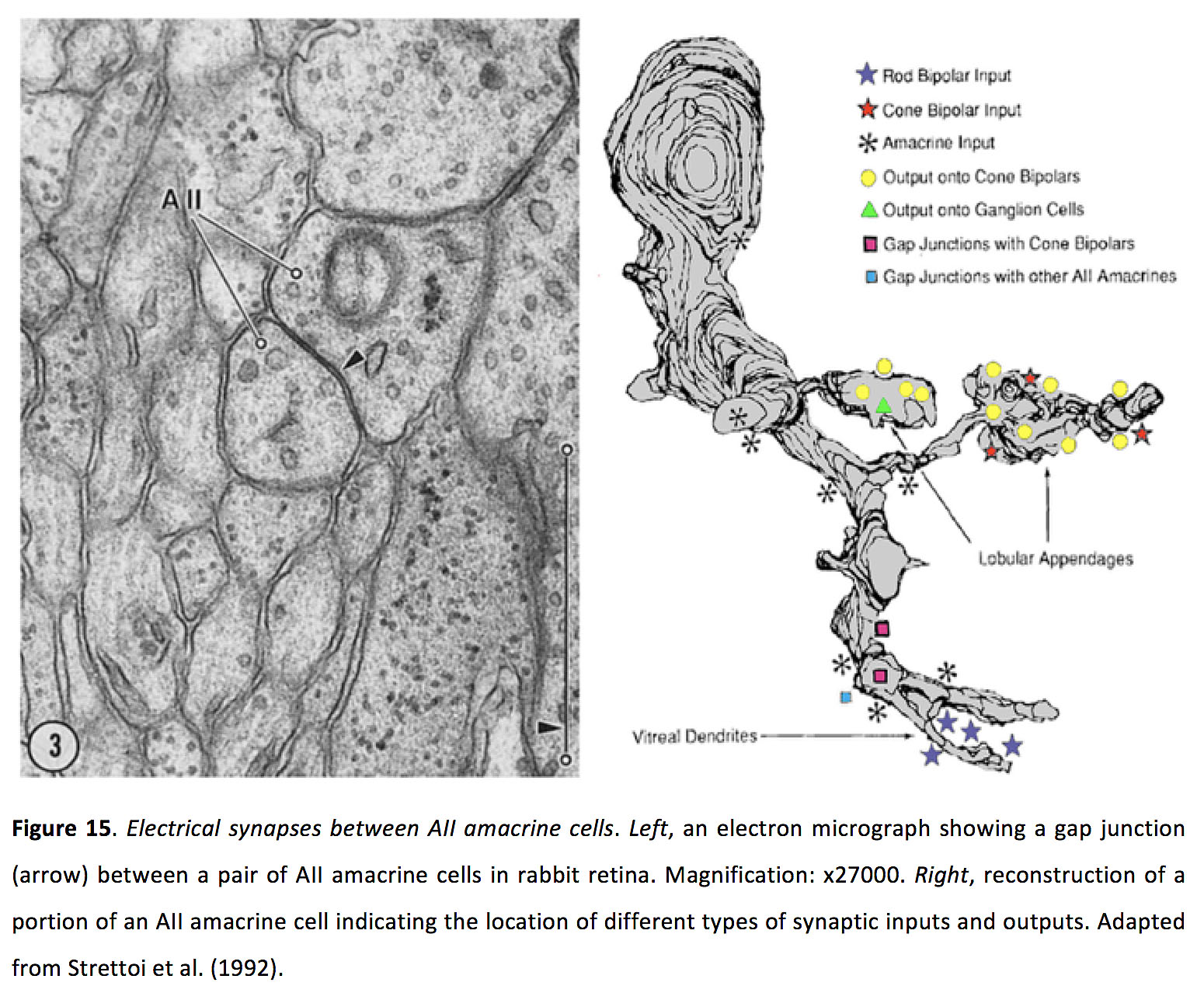 图15.A之间的电突触二世无长突细胞.左图示:兔视网膜上的一对无分泌细胞之间的缝隙连接(箭头)。放大:x27000。正确的,重建的一部分全aiamacrine细胞,表明不同类型的突触输入和输出的位置。改编自Strettoi et al.(1992)(69)。
图15.A之间的电突触二世无长突细胞.左图示:兔视网膜上的一对无分泌细胞之间的缝隙连接(箭头)。放大:x27000。正确的,重建的一部分全aiamacrine细胞,表明不同类型的突触输入和输出的位置。改编自Strettoi et al.(1992)(69)。
所有的无分泌细胞都与ON锥双极细胞与Cx36形成间隙连接。aiamacrine细胞间的耦合是双向对称的,结电导为~700 pS,耦合系数为~ 0.3 (图16)(78)。
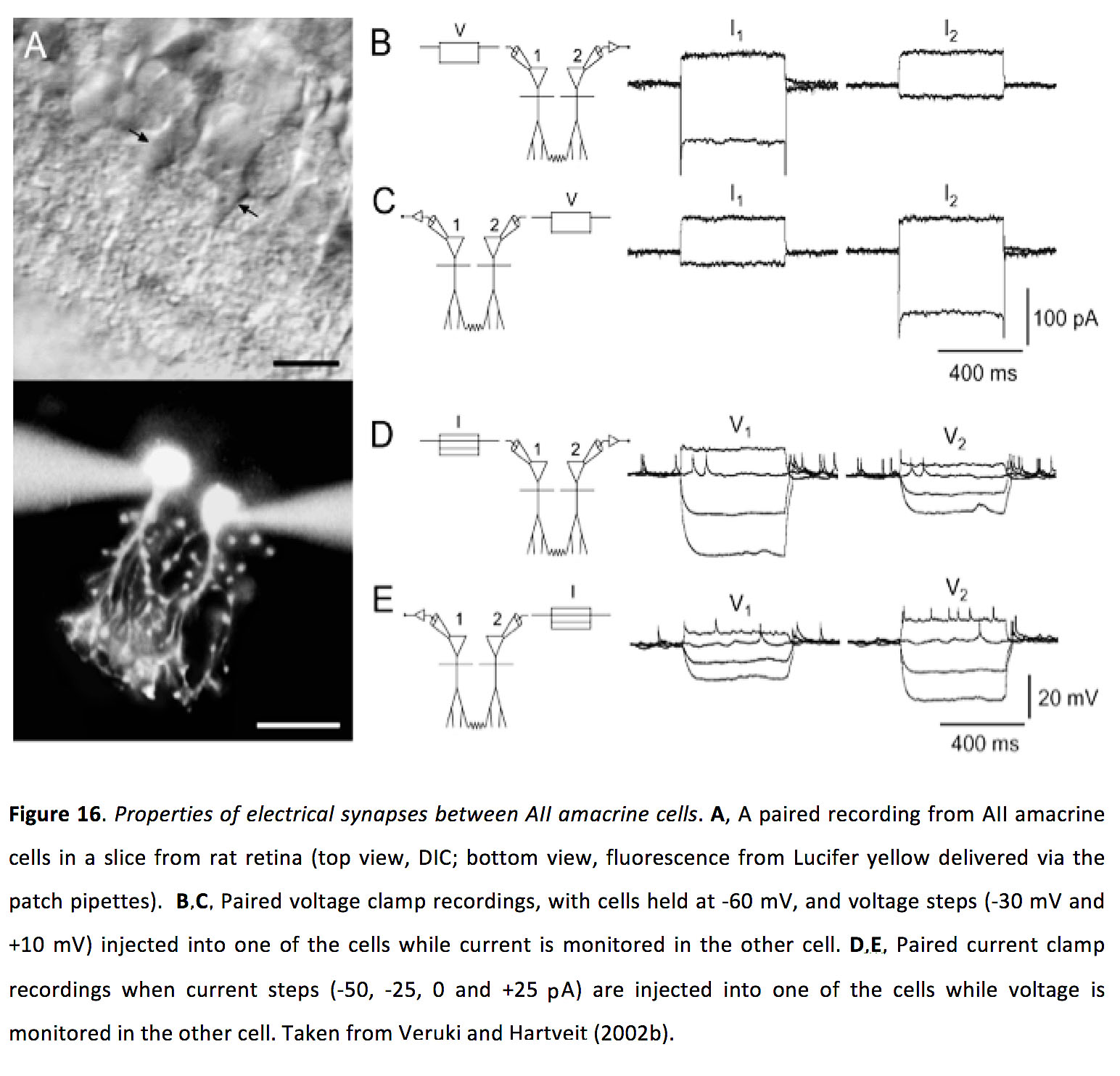 图16.A二世无长突细胞.一个大鼠视网膜切片AIIamacrine细胞的配对记录(俯视图,DIC;底部视图,荧光从路西法黄色输送通过斑块移液管)。B,C配对电压钳记录,电池保持在-60 mV,电压阶跃(-30 mV和+10 mV)注入其中一个电池,同时在另一个电池中监测电流。D,E当电流步进(-50,-25,0和+25 pA)注入其中一个电池,同时在另一个电池中监测电压时,配对电流钳记录。取自Veruki和Hartveit (2002b)(78)。
图16.A二世无长突细胞.一个大鼠视网膜切片AIIamacrine细胞的配对记录(俯视图,DIC;底部视图,荧光从路西法黄色输送通过斑块移液管)。B,C配对电压钳记录,电池保持在-60 mV,电压阶跃(-30 mV和+10 mV)注入其中一个电池,同时在另一个电池中监测电流。D,E当电流步进(-50,-25,0和+25 pA)注入其中一个电池,同时在另一个电池中监测电压时,配对电流钳记录。取自Veruki和Hartveit (2002b)(78)。
其他无分泌细胞类型与无分泌细胞或神经节细胞形成间隙连接,其电突触由Cx36或Cx45组成(96,97)。nNOS-2无分泌细胞是小鼠视网膜中一氧化氮的主要来源,已被证明与似乎含有缝隙连接的Cx45广泛连接,并显示双向和对称的电导,其中一个电导模型非常大(4.4 nS,图17)(98)。测量的nNOS-2无分泌细胞间隙连接的耦合系数为~0.08(98)。
 图17.小鼠视网膜中nNOS-2无腺细胞间的示踪耦合.一个在视野中心注射神经生物素(绿色)的单个无腺细胞进行示踪耦合。B(前),在间隙连接阻滞剂甲氯胺酸存在的情况下,向nNOS-2无分泌细胞注射神经生物素,导致相邻细胞没有示踪剂偶联。红色和粉红色细胞分别表示神经节细胞层(GCL,粉红色)和内核层(INL,红色)附近的nNOS-2无分泌细胞的位置。B(底),充满神经生物素(绿色)的nNOS-2无分泌细胞的侧面投影,与乙酰转移胆碱(ChAT)共染色,该细胞标记星爆无分泌细胞,并有助于勾画出内部丛状层。C,急性视网膜准备的2光子图像显示Alexa Fluor 488(绿色),通过贴片移液管传递到单个nNOS-2无液细胞,可以传播到整个耦合网络。蓝色代表tdTomato转基因标记的nNOS-2大腺细胞。摘自Jacoby等人(2018)(98)。
图17.小鼠视网膜中nNOS-2无腺细胞间的示踪耦合.一个在视野中心注射神经生物素(绿色)的单个无腺细胞进行示踪耦合。B(前),在间隙连接阻滞剂甲氯胺酸存在的情况下,向nNOS-2无分泌细胞注射神经生物素,导致相邻细胞没有示踪剂偶联。红色和粉红色细胞分别表示神经节细胞层(GCL,粉红色)和内核层(INL,红色)附近的nNOS-2无分泌细胞的位置。B(底),充满神经生物素(绿色)的nNOS-2无分泌细胞的侧面投影,与乙酰转移胆碱(ChAT)共染色,该细胞标记星爆无分泌细胞,并有助于勾画出内部丛状层。C,急性视网膜准备的2光子图像显示Alexa Fluor 488(绿色),通过贴片移液管传递到单个nNOS-2无液细胞,可以传播到整个耦合网络。蓝色代表tdTomato转基因标记的nNOS-2大腺细胞。摘自Jacoby等人(2018)(98)。
光适应状态似乎强烈控制无分泌细胞的耦合强度(94)。与水平细胞相似,示踪剂偶联实验表明,所有的无尾细胞在非常昏暗和明亮的条件下偶联极小,而在中等光照条件下偶联强烈(图18)(94)。
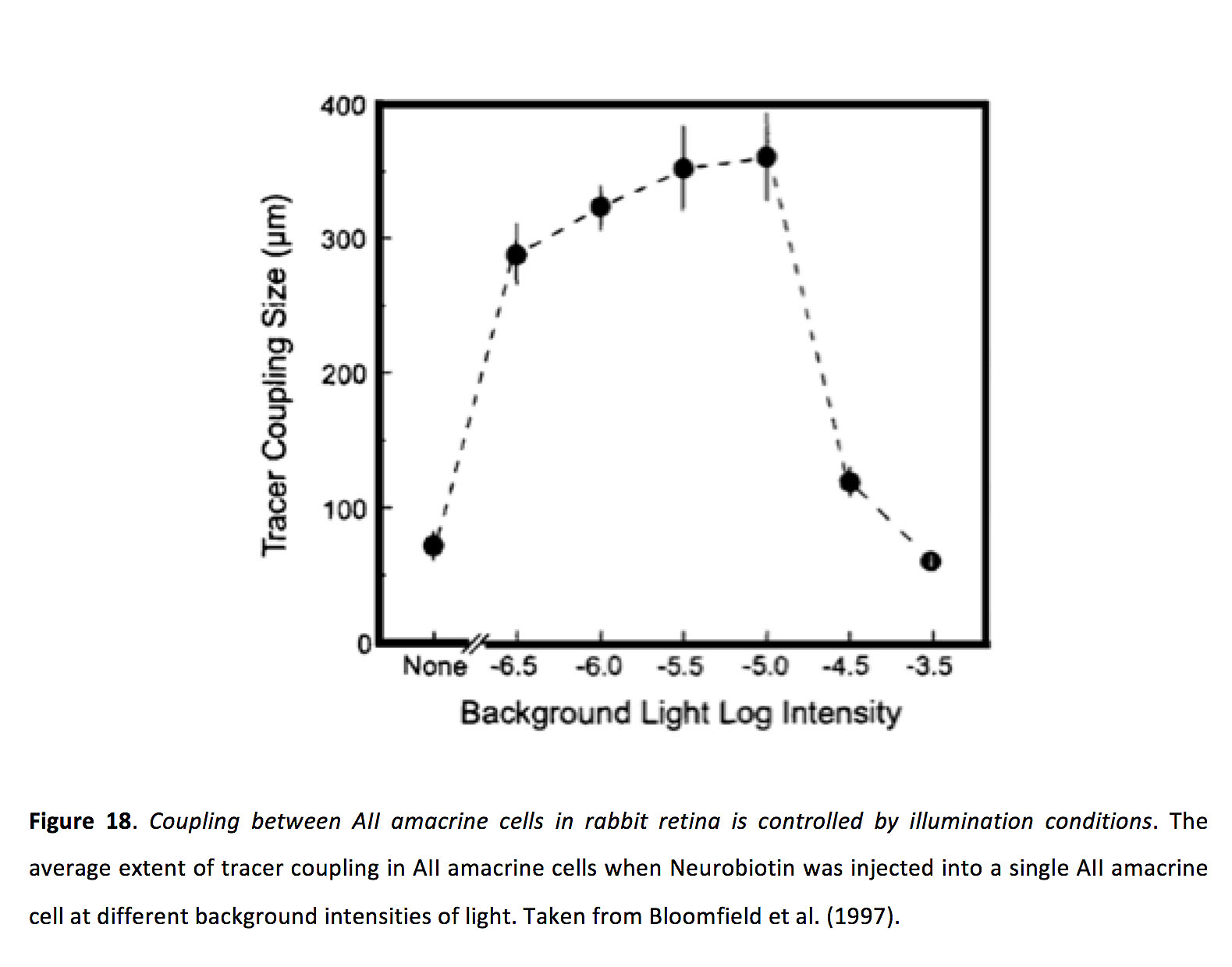 图18.之间的耦合兔视网膜上的无分泌细胞受光照条件的控制.在不同的光背景强度下,将神经生物素注入到单个腺分泌细胞中,示踪剂偶联的平均程度。摘自Bloomfield等人(1997)(200)。
图18.之间的耦合兔视网膜上的无分泌细胞受光照条件的控制.在不同的光背景强度下,将神经生物素注入到单个腺分泌细胞中,示踪剂偶联的平均程度。摘自Bloomfield等人(1997)(200)。
这种耦合强度的变化似乎强烈影响感受野的大小,因为所有无分泌细胞的感受野在光敏状态下可能比树突状野大2-3倍(94)。然而,耦合强度和感受野大小之间的直接关系似乎并不适用于所有的无分泌细胞类型。例如,不像所有的无分泌细胞,与其他一些类型的无分泌细胞的合作并没有表明光适应对感受区大小的强烈影响,尽管事实上示踪剂耦合实验显示示踪剂耦合模式的光适应依赖变化(图19)(94)。为什么间隙结耦合对感受野大小的影响在所有无分泌细胞类型中并不一致还不清楚,尽管这可能与某些无分泌细胞类型的树突似乎执行局部计算有关(99),因此可能不需要感受野大小的全局变化来改变结电导对电路功能产生影响。光适应对无分泌细胞偶联的差异的另一种可能是多巴胺调节的差异。事实上,A8无分泌细胞的间隙连接偶联,通过Cx36间隙连接偶联到ON和OFF双极细胞,似乎不受多巴胺受体D的调节1激活,与所有无分泌细胞相比(100)。最后,在小鼠nNOS-2无分泌细胞中,光激活与耦合状态之间存在显著的自我调节。nNOS-2细胞在适应黑暗的视网膜中广泛电偶联,而在光激活时,它们产生一氧化氮,而一氧化氮反过来又使网络解耦(98)。这种光激活解耦是否会影响感受野的大小还有待测试。
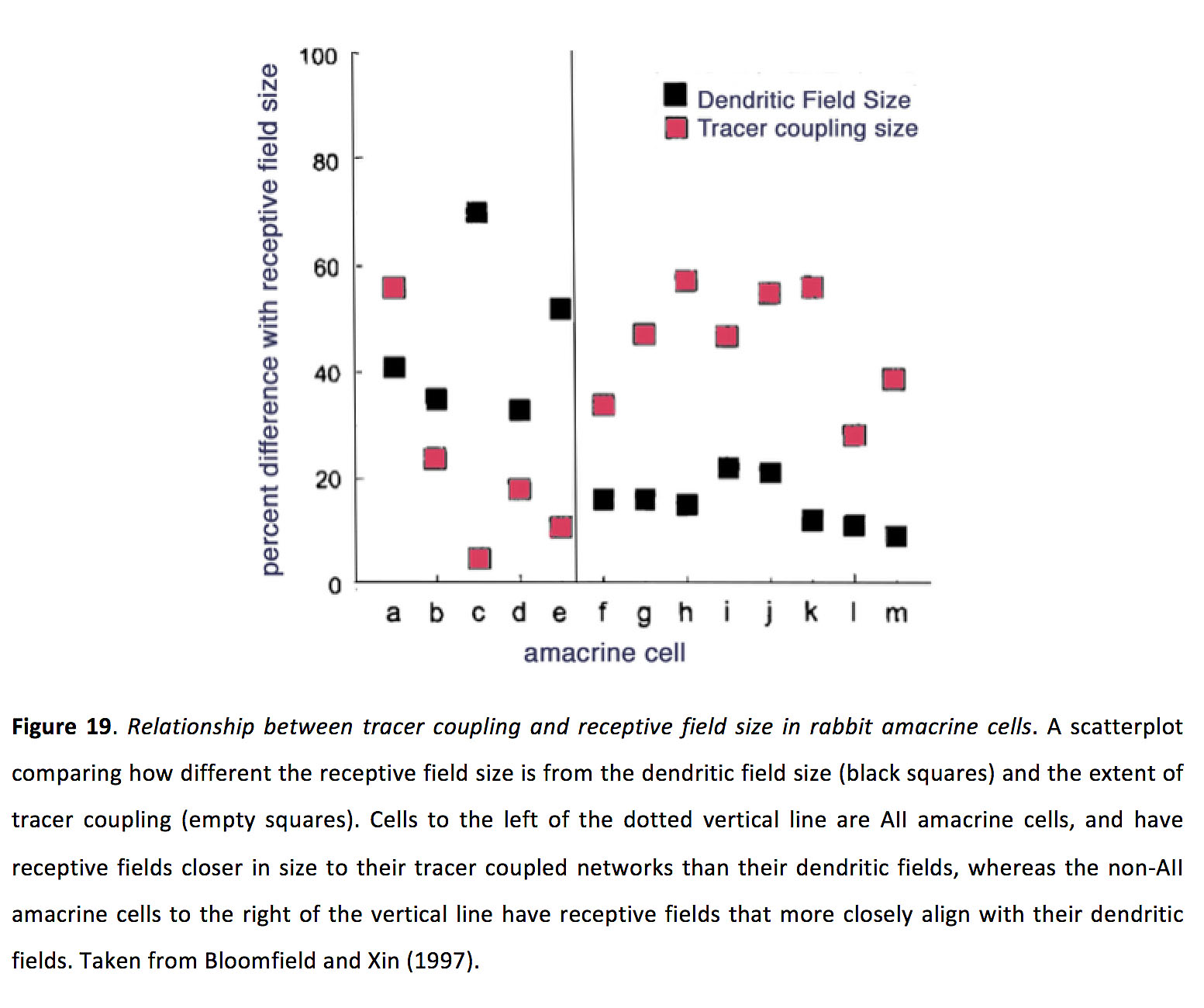
图19.兔大分泌细胞示踪剂偶联与感受野大小的关系.一个散点图,比较感受野大小与树突状野大小(黑色方块)和示踪剂耦合程度(粉色方块)的差异。虚线左侧的细胞均为无分泌细胞,其感受野与示踪耦合网络的距离比树突场的距离更近,而垂直线右侧的非无分泌细胞的感受野与树突场的距离更近。摘自Bloomfield和Xin(1997)(200)。
神经节细胞
神经节细胞有超过30种类型(101-103),从双极细胞接收兴奋性谷氨酸能输入,从无分泌细胞接收抑制性gaba能和甘氨酸能输入,并通过视神经将信号发送到多个更高的视觉区域(6,7)(参见Webvision章节神经节细胞的视觉反应).神经节细胞也与无分泌细胞和其他神经节细胞形成树突间隙连接(图20)(91, 95, 104-108)。
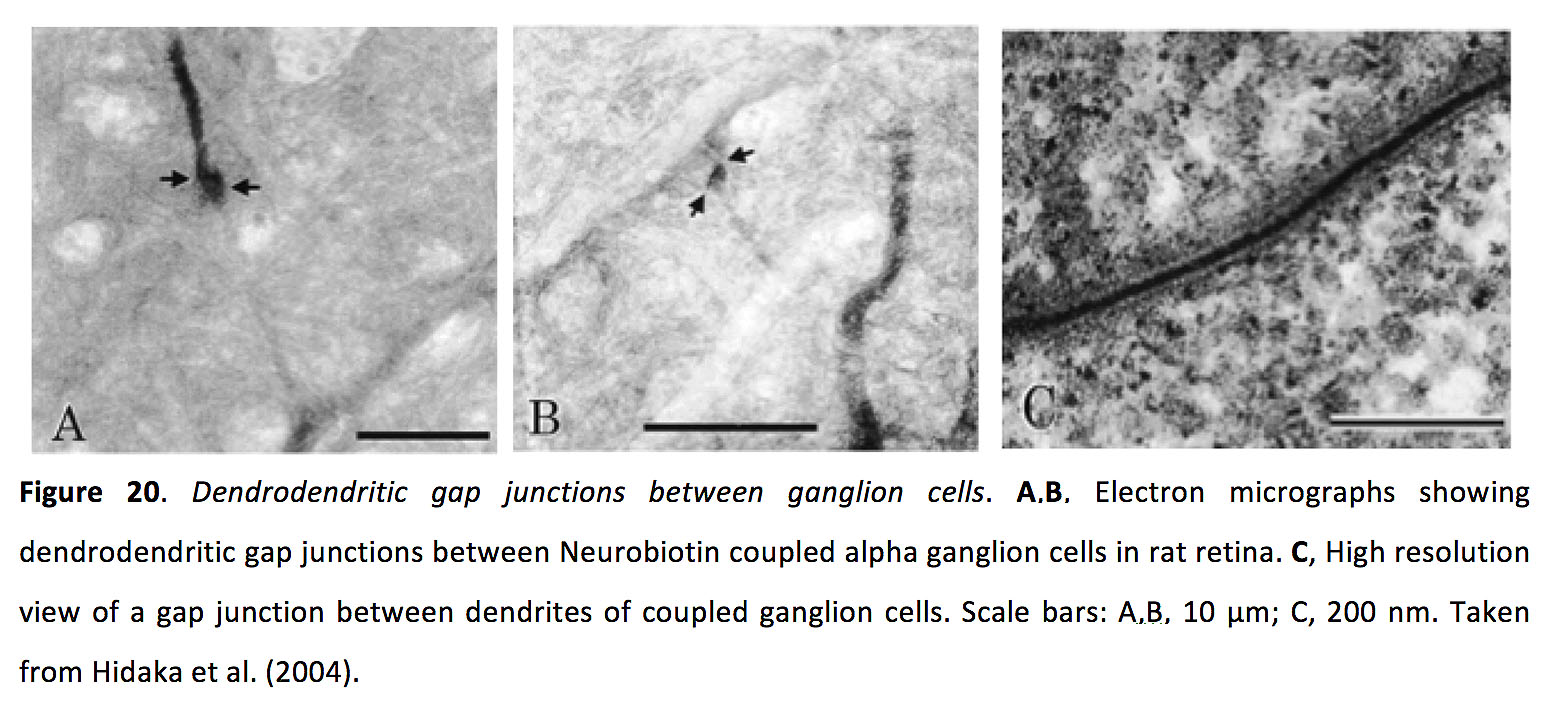 图20.神经节细胞之间的树突间隙连接.一个,B电子显微镜显示大鼠视网膜神经生物素偶联α神经节细胞之间的树突间隙连接。C,偶联神经节细胞树突间间隙连接的高分辨率视图。比例尺:A、B、10µm;C 200海里。取自Hidaka et al.(2004)(106)。
图20.神经节细胞之间的树突间隙连接.一个,B电子显微镜显示大鼠视网膜神经生物素偶联α神经节细胞之间的树突间隙连接。C,偶联神经节细胞树突间间隙连接的高分辨率视图。比例尺:A、B、10µm;C 200海里。取自Hidaka et al.(2004)(106)。
神经节细胞间隙连接通常由连接蛋白36 (图21)(96,109 -111),尽管有些包含连接蛋白45(109),也可能包含连接蛋白30.2(109)。
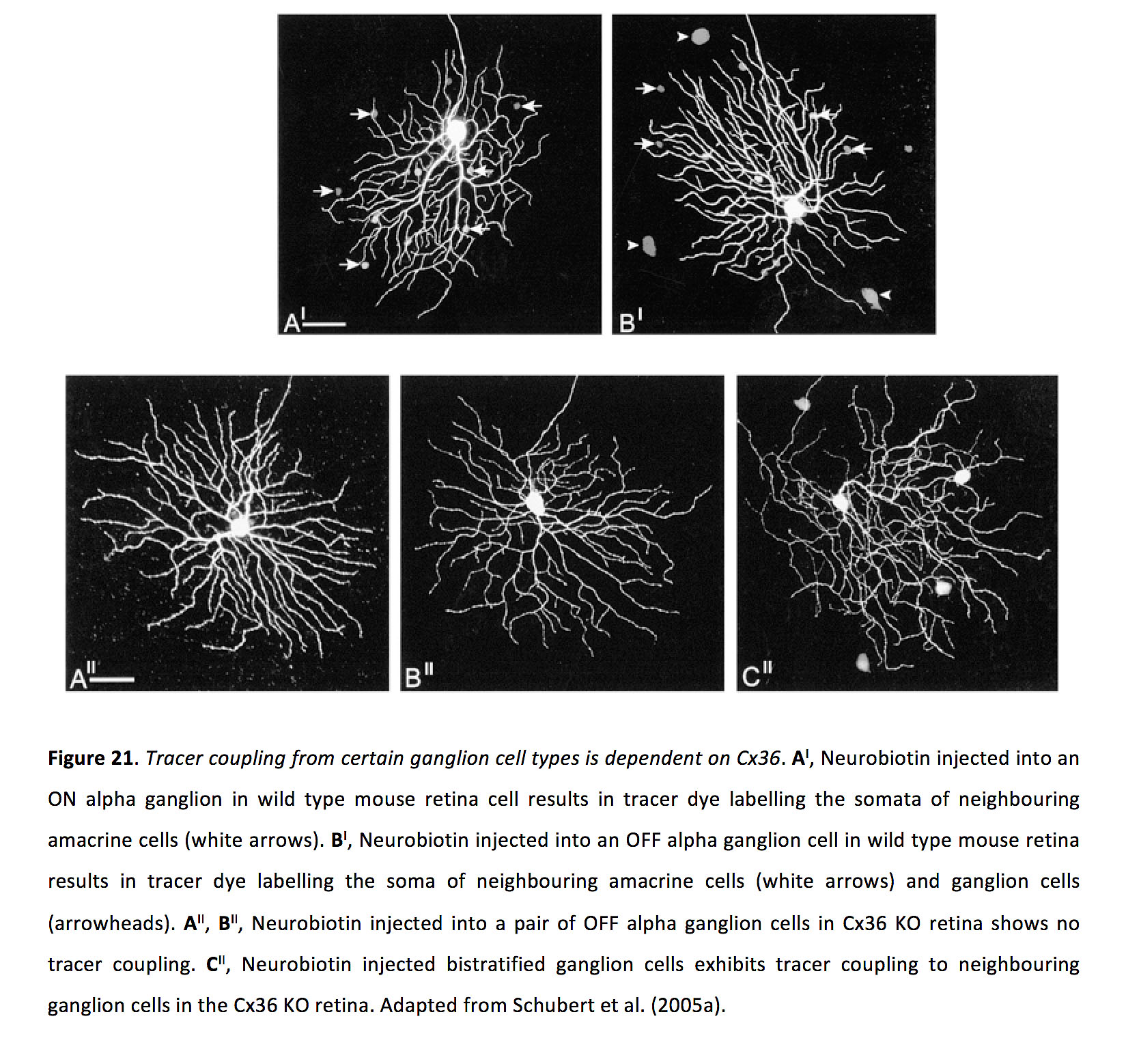 图21.某些神经节细胞类型的示踪剂耦合依赖于Cx36.一个我在野生型小鼠视网膜细胞中,将神经生物素注射到ON α神经节会产生示踪染料,标记邻近的无突细胞的体细胞(白色箭头)。B我在野生型小鼠视网膜的OFF α神经节细胞中注入神经生物素,结果示踪染料标记了邻近的无分泌细胞(白色箭头)和神经节细胞(箭头)的体细胞。一个我我,B我我将神经生物素注射到cx36ko视网膜OFF α神经节细胞中,未见示踪剂偶联。C我我在Cx36 KO视网膜中,神经生物素注射到双层神经节细胞显示示踪剂与邻近的神经节细胞偶联。改编自舒伯特等人(2005a)(201)。
图21.某些神经节细胞类型的示踪剂耦合依赖于Cx36.一个我在野生型小鼠视网膜细胞中,将神经生物素注射到ON α神经节会产生示踪染料,标记邻近的无突细胞的体细胞(白色箭头)。B我在野生型小鼠视网膜的OFF α神经节细胞中注入神经生物素,结果示踪染料标记了邻近的无分泌细胞(白色箭头)和神经节细胞(箭头)的体细胞。一个我我,B我我将神经生物素注射到cx36ko视网膜OFF α神经节细胞中,未见示踪剂偶联。C我我在Cx36 KO视网膜中,神经生物素注射到双层神经节细胞显示示踪剂与邻近的神经节细胞偶联。改编自舒伯特等人(2005a)(201)。
神经节细胞到神经节细胞的偶联似乎是完全同源的,不同类型的神经节细胞之间没有偶联(95,105,112,113),尽管可能有例外(114)。此外,不同的神经节细胞亚型具有独特的耦合模式(图22)(95)。尽管示踪剂偶联工作表明特定的神经节细胞与特定类型的无泡细胞形成缝隙连接,但对神经节细胞-无泡细胞偶联知之甚少(图22)(95)。
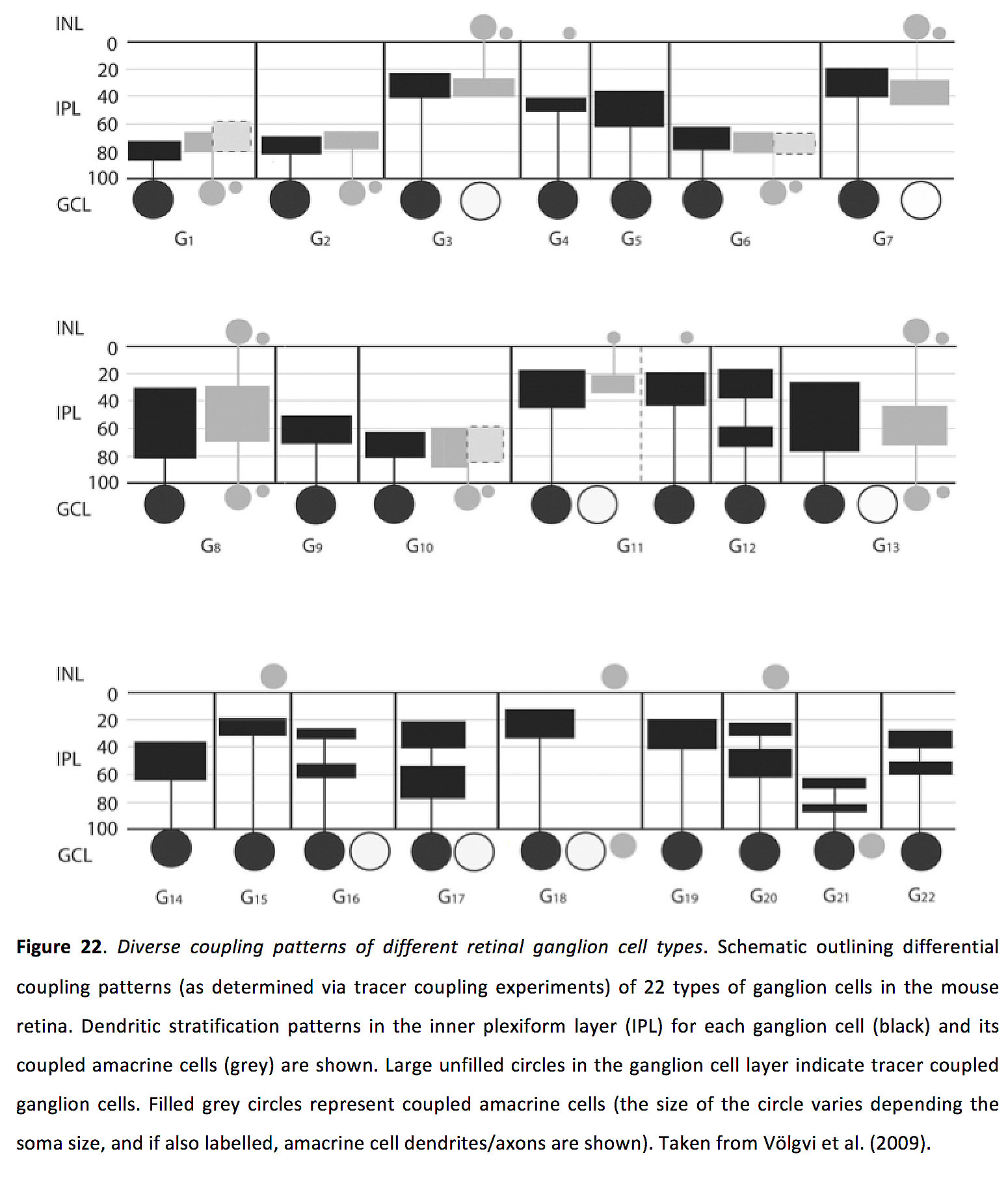 图22.不同视网膜神经节细胞类型的不同耦合模式.图示小鼠视网膜22种神经节细胞的差异耦合模式(通过示踪耦合实验确定)。每个神经节细胞(黑色)和其偶联的无分泌细胞(灰色)内丛状层(IPL)的树突状分层模式显示。神经节细胞层中未填充的大圆圈表示示踪剂偶联的神经节细胞。填充的灰色圆圈表示偶联的无分泌细胞(圆圈的大小取决于体细胞的大小,如果也标记,则显示无分泌细胞的树突/轴突)。摘自Völgyi等(2009)(95)。
图22.不同视网膜神经节细胞类型的不同耦合模式.图示小鼠视网膜22种神经节细胞的差异耦合模式(通过示踪耦合实验确定)。每个神经节细胞(黑色)和其偶联的无分泌细胞(灰色)内丛状层(IPL)的树突状分层模式显示。神经节细胞层中未填充的大圆圈表示示踪剂偶联的神经节细胞。填充的灰色圆圈表示偶联的无分泌细胞(圆圈的大小取决于体细胞的大小,如果也标记,则显示无分泌细胞的树突/轴突)。摘自Völgyi等(2009)(95)。
神经节细胞之间的间隙连接是双向的,具有约1ns的对称连接电导和约0.15 (图23)(106, 113)。
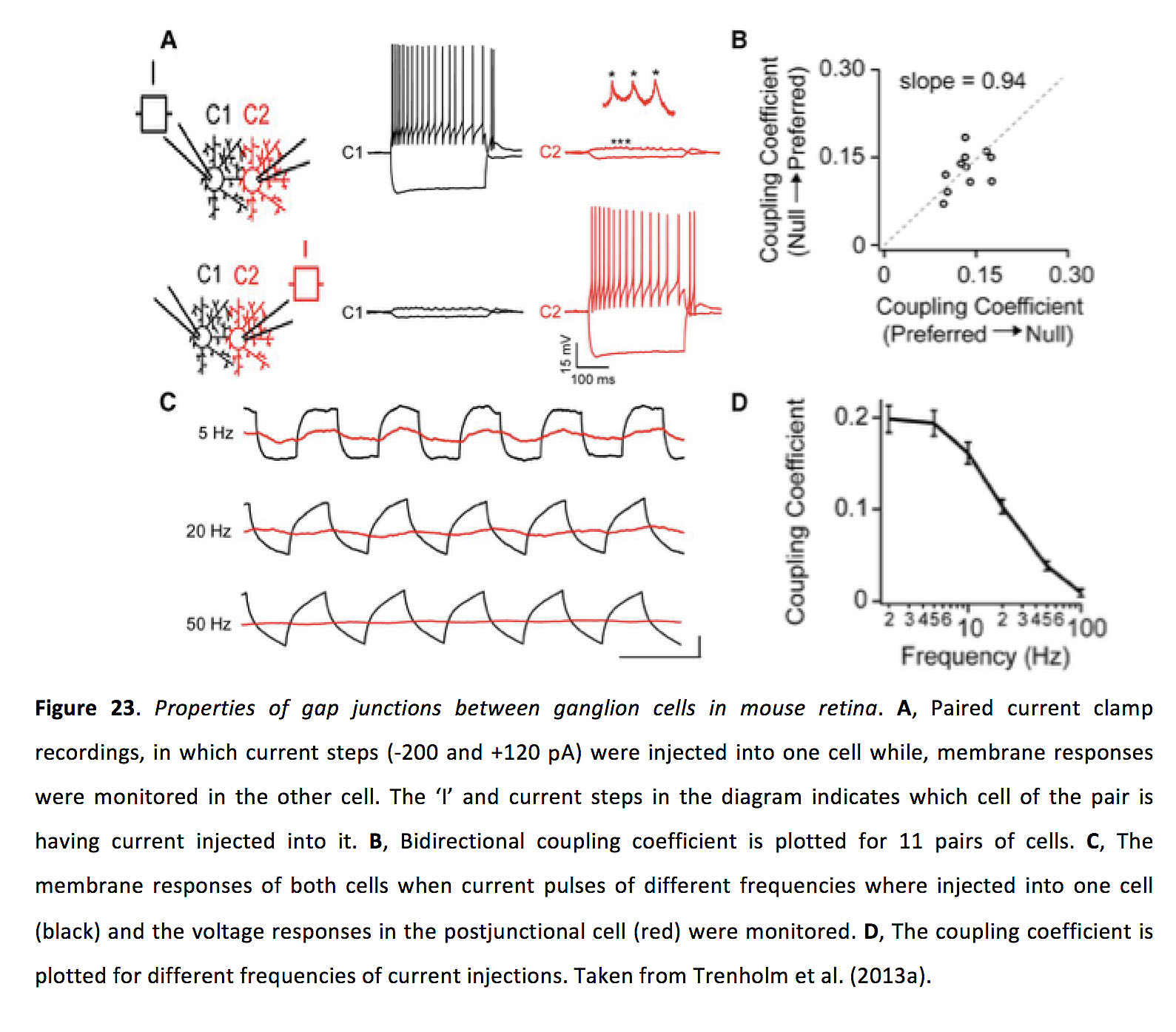 图23.小鼠视网膜神经节细胞间缝隙连接的特性.一个配对电流钳记录,将电流步骤(-200和+120 pA)注入一个细胞,同时监测另一个细胞的膜反应。图中的“I”和电流步骤表示电流注入到哪个细胞中。B,绘制了11对单元的双向耦合系数。C当不同频率的电流脉冲注入一个细胞(黑色)和连接后细胞(红色)时,观察两个细胞的膜响应。D,绘制了不同电流注入频率下的耦合系数。取自Trenholm等人(2013a)(113)。
图23.小鼠视网膜神经节细胞间缝隙连接的特性.一个配对电流钳记录,将电流步骤(-200和+120 pA)注入一个细胞,同时监测另一个细胞的膜反应。图中的“I”和电流步骤表示电流注入到哪个细胞中。B,绘制了11对单元的双向耦合系数。C当不同频率的电流脉冲注入一个细胞(黑色)和连接后细胞(红色)时,观察两个细胞的膜响应。D,绘制了不同电流注入频率下的耦合系数。取自Trenholm等人(2013a)(113)。
在神经节细胞中,光适应似乎控制偶联强度,在黑暗适应状态下偶联减弱,而在光适应状态下偶联增强(至少兔视网膜OFF α细胞是这样的(115);虽然这可能不是所有神经节细胞类型的情况(111))。有趣的是,虽然耦合强度的增加似乎并没有改变经典(峰值)感受野的大小(115,116),但来自邻近细胞的缝隙连接输入赋予电耦合神经节细胞广泛的阈下兴奋周围感受野(113)。此外,对于许多神经节细胞来说,在光适应条件下,感受野的大小实际上减小了(117-119),这可能是由于水平和/或无分泌细胞产生的抑抑性周围反应的加强。例如,研究发现,对于小鼠ON和OFF α样神经节细胞(59)和定向选择性神经节细胞(60),神经节细胞中的包围抑制是由宽视野无分泌细胞提供的,而且这种包围抑制似乎只有在激活视锥的光强下才被激活。在这种明亮的光线下,无分泌细胞介导的包围抑制被认为依赖于宽场无分泌细胞和on锥双极细胞之间的电耦合。敲除Cx36可以消除强光引起的α样神经节细胞感受野大小的减少(59)。因此,神经节细胞中的经典感受野的大小似乎最受无分泌细胞介导的周围抑制的控制:最近的一项研究表明,水平细胞可能只对神经节细胞的周围抑制贡献约15%(58)。
关于神经节细胞-大泡细胞耦合的特性,我们所知甚少。尽管如此,缝隙连接的强度似乎足够强,以至于注入一个神经节细胞的电流可以通过缝隙连接进入一个无分泌细胞,然后通过额外的缝隙连接进入附近的神经节细胞,在间接耦合的神经节细胞中产生可测量的反应(图24)(120)。
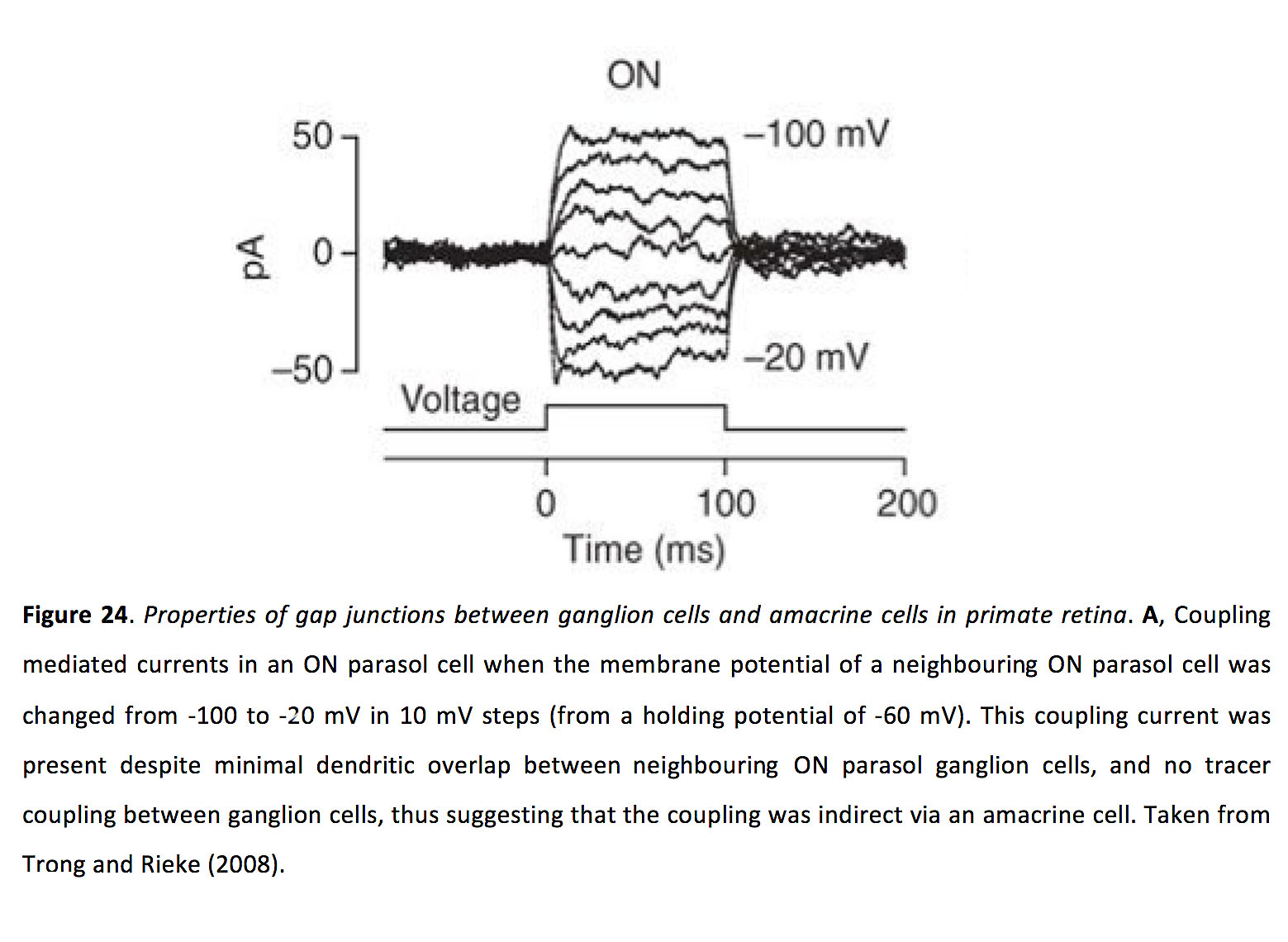 图24.灵长类动物视网膜神经节细胞与无分泌细胞间隙连接的特性.一个当相邻ON阳伞细胞的膜电位在10 mV步骤内从-100 mV变为-20 mV时(从-60 mV的保持电位),ON阳伞细胞内的耦合介导电流。尽管相邻的ON阳伞神经节细胞之间的树突重叠极小,而且神经节细胞之间没有示踪剂偶联,但这种偶联电流仍然存在,因此表明这种偶联是通过无分泌细胞间接产生的。摘自Trong和Rieke(2008)(120)。
图24.灵长类动物视网膜神经节细胞与无分泌细胞间隙连接的特性.一个当相邻ON阳伞细胞的膜电位在10 mV步骤内从-100 mV变为-20 mV时(从-60 mV的保持电位),ON阳伞细胞内的耦合介导电流。尽管相邻的ON阳伞神经节细胞之间的树突重叠极小,而且神经节细胞之间没有示踪剂偶联,但这种偶联电流仍然存在,因此表明这种偶联是通过无分泌细胞间接产生的。摘自Trong和Rieke(2008)(120)。
缝隙连接和暗痣视觉
在暗(暗)光条件下,视杆细胞是活跃的光感受器,间隙连接对于视杆介导的信号通过视网膜至关重要。在经典的杆状通路中,杆状细胞与杆状双极细胞形成谷氨酸突触,杆状双极细胞又与所有无分泌细胞形成谷氨酸突触。这些无分泌细胞通过与ON锥双极细胞的间隙连接,将杆状细胞产生的信号转移到ON锥双极细胞。此外,所有的无分泌细胞也与OFF双极细胞产生甘氨酸突触(图25一个)(64, 79, 80, 121-124)。在第二种途径中,视杆细胞的信号通过视杆细胞电突触直接传递给视锥细胞,并由此传递到视网膜内部(图25 B)(36, 122, 125, 126)。在第三种途径中,一些杆似乎与OFF双极细胞形成谷氨酸突触,并且假设电耦合杆之间的信号聚集可以增加该途径的敏感性(图25 C)(17, 127, 128)。
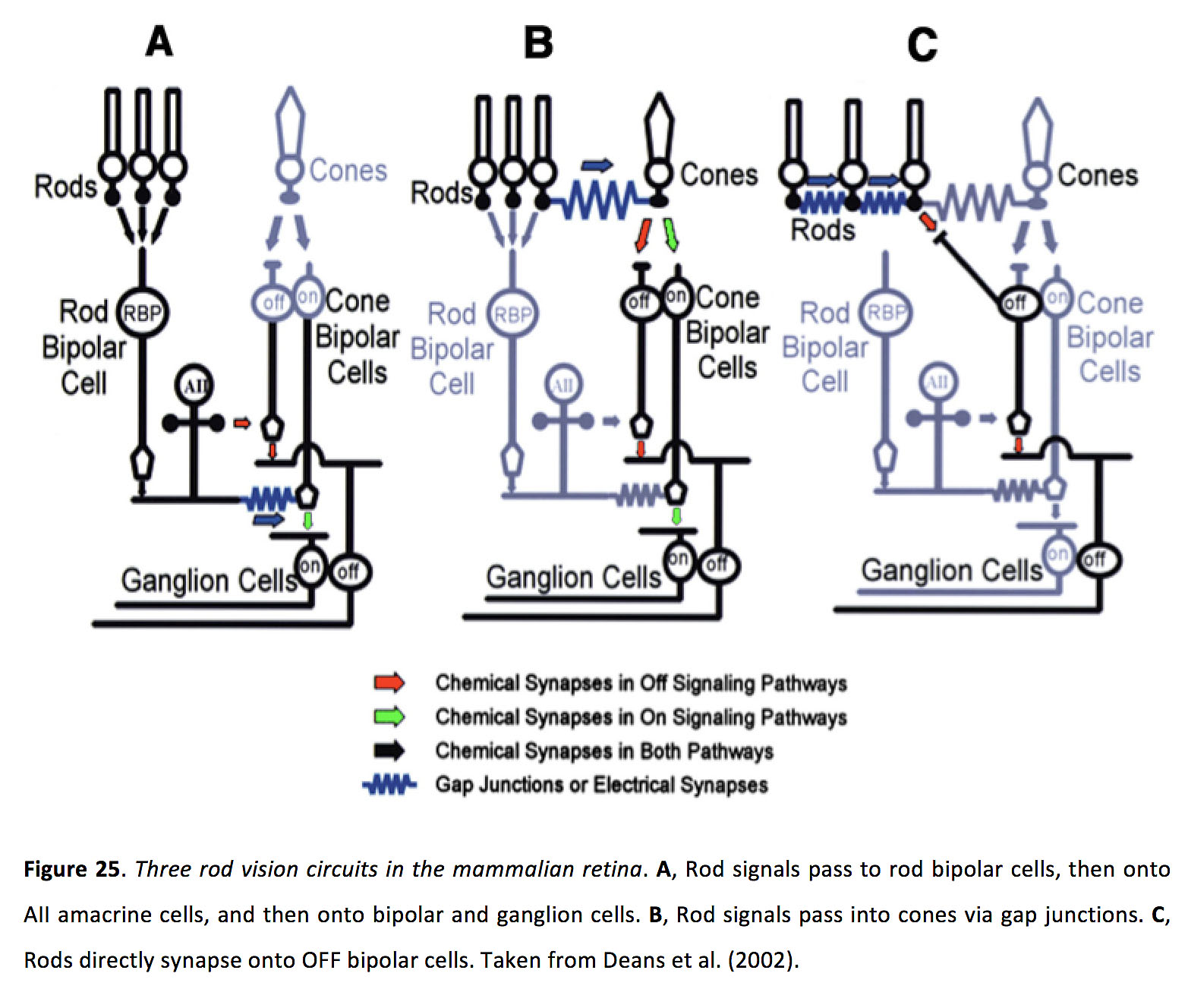 图25.哺乳动物视网膜中的三杆视觉回路.一个杆状信号通过杆状双极细胞,然后通过所有的无分泌细胞,再通过双极细胞和神经节细胞。B,杆状信号通过缝隙结进入锥。C,棒直接突触到OFF双极细胞。摘自迪恩斯等人(2002)(70)。
图25.哺乳动物视网膜中的三杆视觉回路.一个杆状信号通过杆状双极细胞,然后通过所有的无分泌细胞,再通过双极细胞和神经节细胞。B,杆状信号通过缝隙结进入锥。C,棒直接突触到OFF双极细胞。摘自迪恩斯等人(2002)(70)。
与杆介导的短视视觉中缝隙连接信号的重要作用一致,敲除Cx36会大大损害视网膜内杆介导的反应(图26)(70, 122, 126)。此外,某些类型的无腺分泌和神经节细胞之间的间隙连接也有助于对昏暗光线的高灵敏度检测(129)。
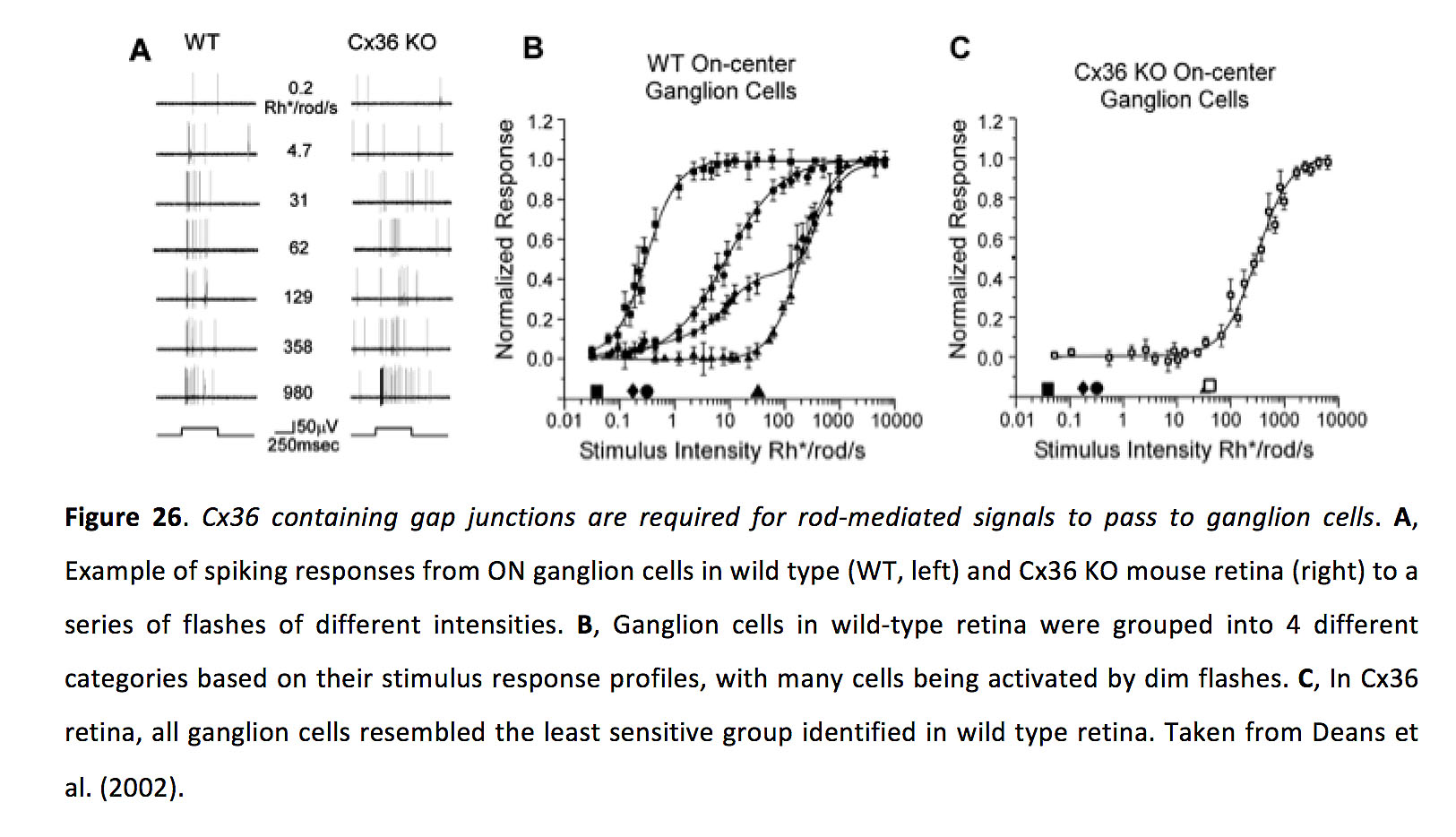 图26.含有间隙连接的Cx36是杆状细胞介导信号传递到神经节细胞所必需的.一个野生型(WT,左)和Cx36 KO小鼠视网膜(右)对一系列不同强度的闪光的ON神经节细胞的脉冲反应的例子。B根据刺激反应谱,野生型视网膜的神经节细胞被分为4类,许多细胞被微弱的闪光激活。C在Cx36视网膜中,所有神经节细胞与野生型视网膜中最不敏感的一组细胞相似。摘自迪恩斯等人(2002)(70)。
图26.含有间隙连接的Cx36是杆状细胞介导信号传递到神经节细胞所必需的.一个野生型(WT,左)和Cx36 KO小鼠视网膜(右)对一系列不同强度的闪光的ON神经节细胞的脉冲反应的例子。B根据刺激反应谱,野生型视网膜的神经节细胞被分为4类,许多细胞被微弱的闪光激活。C在Cx36视网膜中,所有神经节细胞与野生型视网膜中最不敏感的一组细胞相似。摘自迪恩斯等人(2002)(70)。
隙结被认为在棒的信号传递中扮演着另一个重要的角色,它允许棒提高信噪比。光感受器的固有噪声是单个细胞独有的,而间隙结耦合允许光诱发信号共享,从而更有效地检测相对微弱的信号,并减少响应的可变性。但请注意,这些增益可能导致在昏暗条件下失去绝对灵敏度和分辨率(12,13,15,32,130 -132)。缝隙连接在杆状信号传递中的最后一个可能的作用与杆状细胞和杆状双极细胞之间的突触传递的性质有关。在杆状细胞和杆状双极细胞之间似乎存在着显著的信号剪切(即突触传递的饱和),并且已经确定,杆状之间的电耦合可以允许空间受限的视觉刺激通过连接扩散激活多个杆状,而这组被激活的杆状细胞将更有效地激活双极细胞(133)。
缝隙连接和同步
缝隙连接在中枢神经系统中被描述得最好的角色之一是它们可以同步耦合细胞之间的活动(134,135)。对视网膜的经典研究显示,邻近的视网膜神经节细胞表现出显著的尖峰同步(136-139)。随后的研究证实,在许多物种中,电突触负责驱动神经节细胞之间不同类型的精细同步(112,116,140 -143)。
相邻神经节细胞之间的相关性通常根据时间特征分为窄、中或宽(图27)(140, 142, 144)。窄同步发生在±2毫秒内,在0毫秒延迟的交叉相关图中有一个波谷。中等相关性在0毫秒延迟时有一个交叉相关图峰值,并出现在2-10毫秒(144)、~ 25-50毫秒(140)和小于100毫秒(142)之间的某个地方。广义相关在交叉相关图中通常表现为0毫秒的延迟峰值,并在大约50毫秒(144)、大于50毫秒(140)和大于100毫秒(142)的时间尺度上出现,但在on和OFF神经节细胞对之间,广义相关可在0毫秒出现波谷,表明缓慢的反相关(137,138,144)。
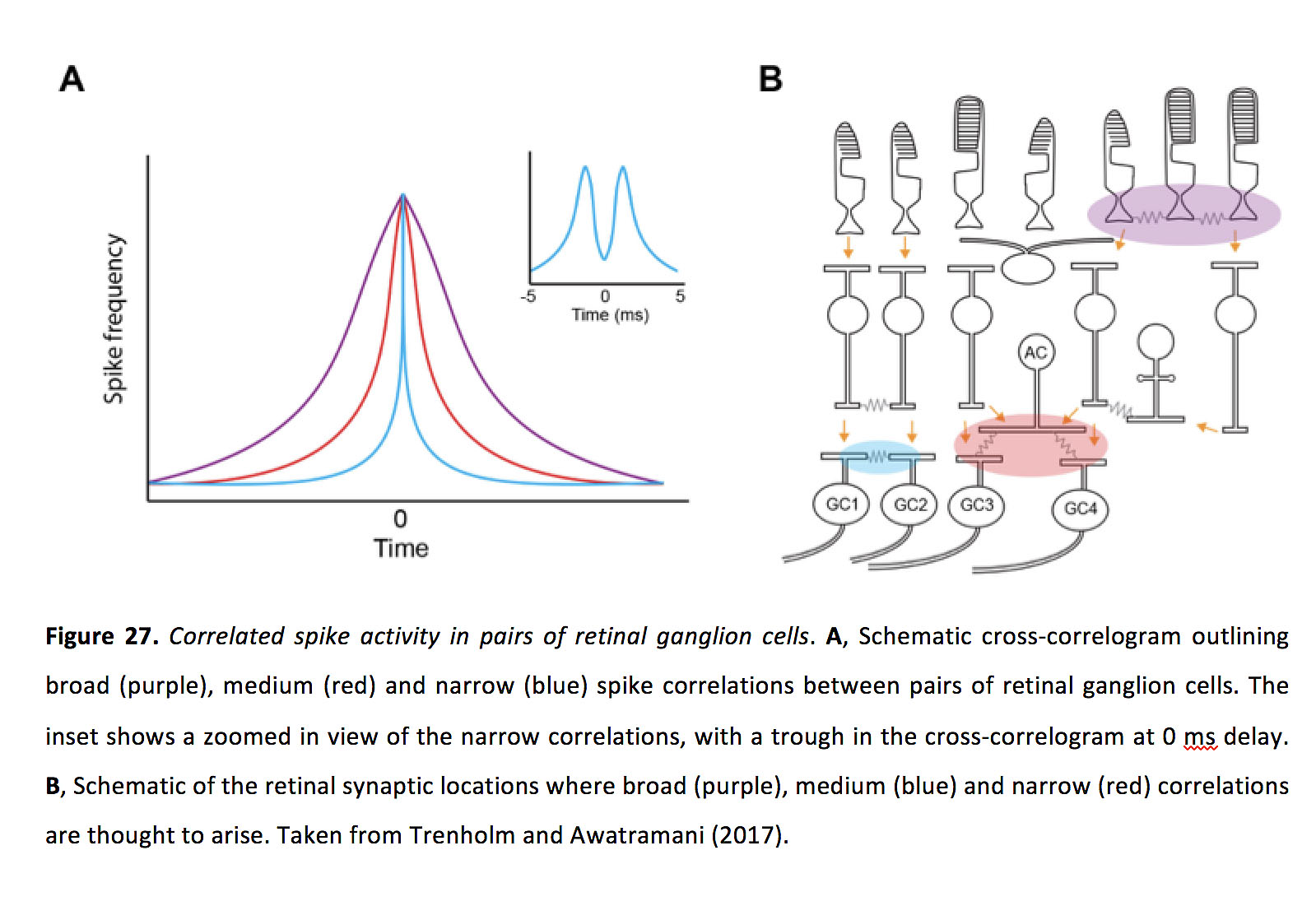 图27所示。对视网膜神经节细胞的相关峰值活性.一个交叉相关图示意图,勾勒出视网膜神经节细胞对之间的宽(紫色)、中(红色)和窄(蓝色)峰相关性。插图显示了窄相关的放大视图,在0毫秒延迟的交叉相关图中有一个波谷。B,宽(紫色)、中(蓝色)和窄(红色)相关的视网膜突触位置示意图。摘自Trenholm and Awatramani(2017)(197)。
图27所示。对视网膜神经节细胞的相关峰值活性.一个交叉相关图示意图,勾勒出视网膜神经节细胞对之间的宽(紫色)、中(红色)和窄(蓝色)峰相关性。插图显示了窄相关的放大视图,在0毫秒延迟的交叉相关图中有一个波谷。B,宽(紫色)、中(蓝色)和窄(红色)相关的视网膜突触位置示意图。摘自Trenholm and Awatramani(2017)(197)。
药理学和基因敲除实验已经进行,以梳理出电和化学突触传递在这些不同类型的神经节细胞相关性中的作用。药理学研究表明,化学突触阻滞剂的混合物不影响狭窄和中等相关性,但缝隙连接阻滞剂的应用会抑制这种相关性(140)。Cx36敲除实验表明,广泛的相关性也可能依赖于间隙结信号(142)。基于这些研究,人们认为广泛的相关性最初产生于光感受器。相反,当对神经节细胞通过无分泌细胞间接耦合(116,140,142,144,145)时,产生中等相关性,而在相互耦合的神经节细胞之间形成狭窄相关性(106,111,112,116,139,140,142,143)。在本节的其余部分,我们主要关注神经节细胞之间的同源耦合和窄相关。
人们认为,当一个神经元发出动作电位时,就会产生窄峰同步,从而使相邻神经元在短时间内产生偏差。然而,由于间隙连接作为低通滤波器,这大大降低了动作电位介导的信号通过连接的振幅(图28)(134, 135, 146),还不清楚为什么如此小的信号可以产生如此强大的同步(120)。
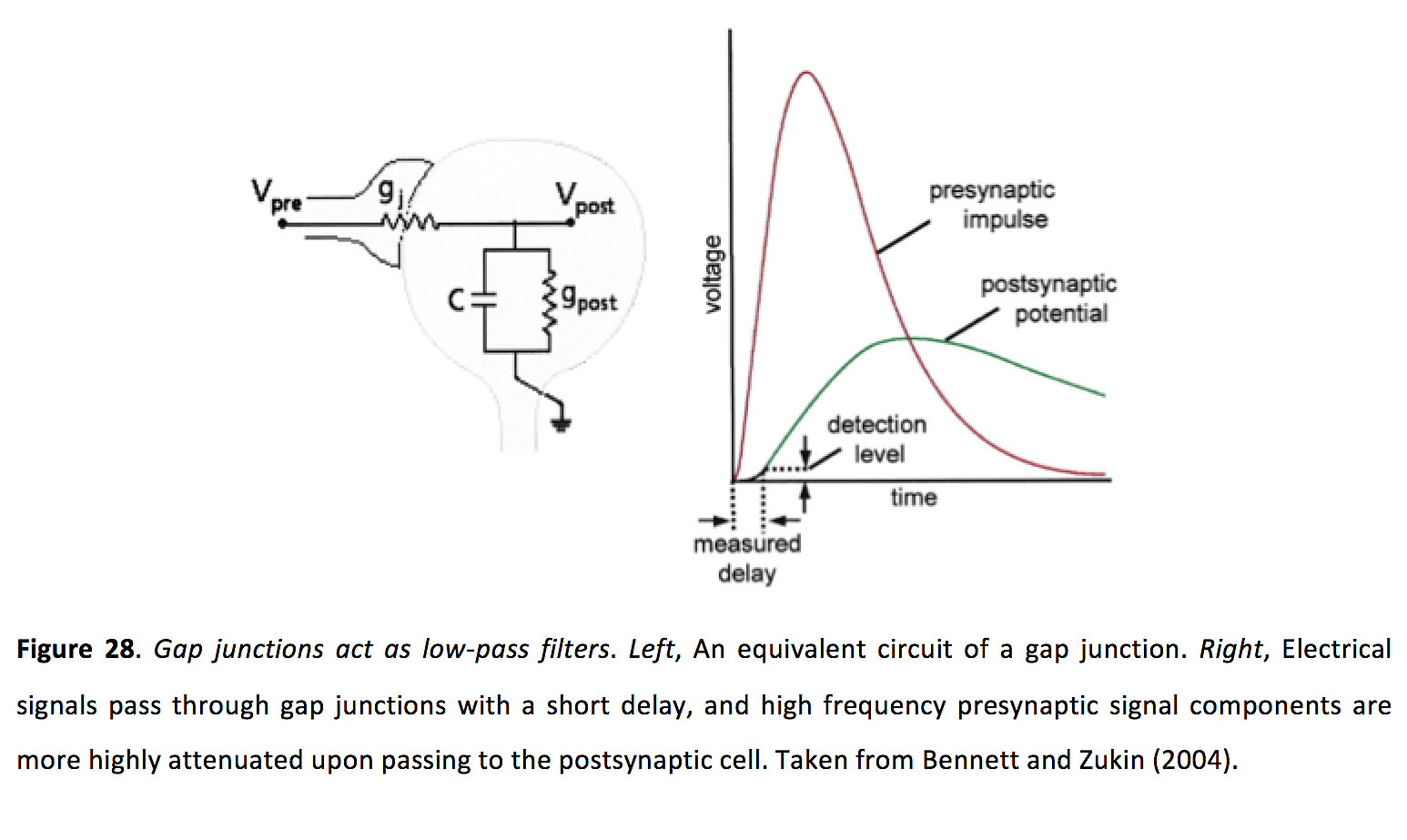 图28.缝隙连接充当低通滤波器.左,间隙结的等效电路。正确的电信号通过间隙连接的延迟较短,而高频的突触前信号成分在传递到突触后细胞时衰减更大。摘自Bennett and Zukin(2004)(146)。
图28.缝隙连接充当低通滤波器.左,间隙结的等效电路。正确的电信号通过间隙连接的延迟较短,而高频的突触前信号成分在传递到突触后细胞时衰减更大。摘自Bennett and Zukin(2004)(146)。
虽然有人认为,一个神经节细胞中的动作电位可以直接驱动相邻的耦合神经节细胞的峰值(112),但更直接的实验证据表明,一个神经节细胞中的动作电位实际上从未驱动耦合神经节细胞的阈上反应(143)。与这一发现相一致的是,该领域的早期工作表明,一个神经节细胞的峰值将直接驱动耦合相邻细胞的峰值,成功率低于5%(139)。此外,神经节细胞的经典感受野不会因间隙连接的存在而显著增加(见112、113、116)。为什么一个神经节细胞的尖刺在直接驱动邻近细胞的尖刺时无效?结果表明,在神经节细胞中,偶联小穗的振幅很小——至少在连接后体细胞用膜片移液管测量时是这样的(图23;~ 1 mV),尽管它们到达时几乎没有延迟(< 1 ms)(113)。因此,神经节细胞之间的耦合类似于中枢神经系统中大多数其他神经元之间的耦合,其中连接前神经元的峰值仅驱动连接后邻近神经元的阈下活动(134)。那么阈下耦合小穗是如何驱动耦合神经节细胞的同步活动的呢?
开关定向选择性(DS)神经节细胞是研究耦合小穗如何通过非线性树突信号驱动同步的理想模型。DS神经节细胞产生Na+-通道依赖,ttx敏感的树突尖峰,高可靠性触发动作电位(147,148)。至少有四种类型的ON-OFF DS神经节细胞,编码基本方向(149)。由于尚不完全清楚的原因,只有喜欢在视野中上方运动(视网膜的腹侧运动)的on - off DS细胞在成年哺乳动物视网膜中表现出同源电耦合,至少在兔子和老鼠的视网膜中是这样(图29)(105, 113, 150)。
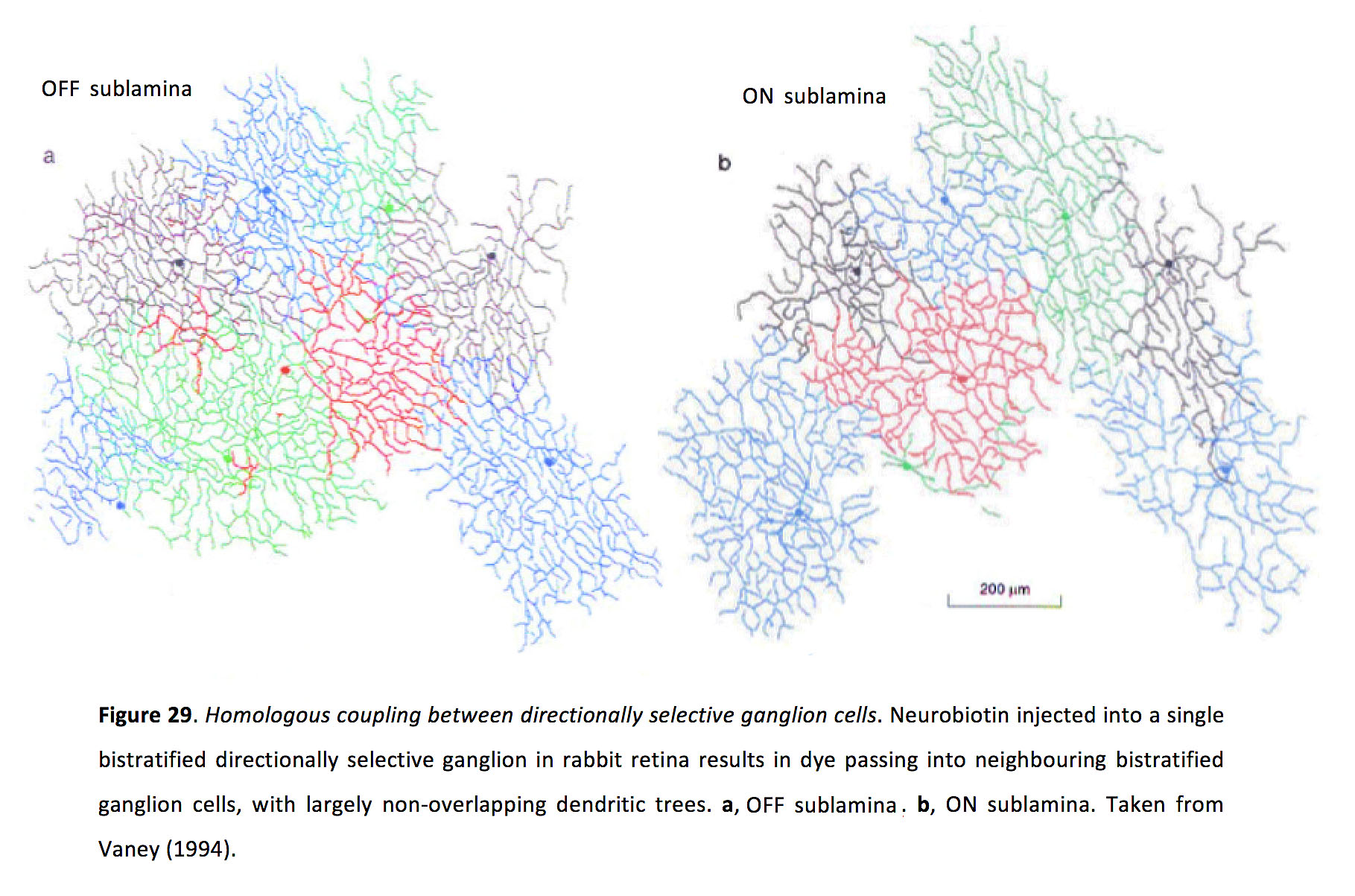 图29.定向选择神经节细胞之间的同源偶联.将神经生物素注入兔视网膜的单个双层定向选择性神经节,可导致染料进入相邻的双层神经节细胞,形成大量不重叠的树突树。一个, sublamina。bsublamina。取自Vaney(1994)(105)。
图29.定向选择神经节细胞之间的同源偶联.将神经生物素注入兔视网膜的单个双层定向选择性神经节,可导致染料进入相邻的双层神经节细胞,形成大量不重叠的树突树。一个, sublamina。bsublamina。取自Vaney(1994)(105)。
有趣的是,当偶联小穗通过在偶联DS神经节细胞的胞体上注入电流(用贴片移液管)模拟时,它们并没有显著地调节动作电位时间,这增加了它们的作用位点在树突中的可能性(143)。此外,耦合的小穗似乎不够大,不能直接启动树突刺。事实证明,要在邻近细胞中驱动脉冲同步,耦合的小脉冲必须与邻近神经节细胞重叠树突中的化学突触输入进行非线性相互作用(143)。在一对相邻的耦合神经节细胞之间接受重叠的共享位置上闪烁的光会驱动显著的峰值相关性,而在两个不同的空间位置同时闪烁两个光点来激活同一对神经节,但没有重叠的化学突触输入,不会驱动显著相关性(图30)(143)。
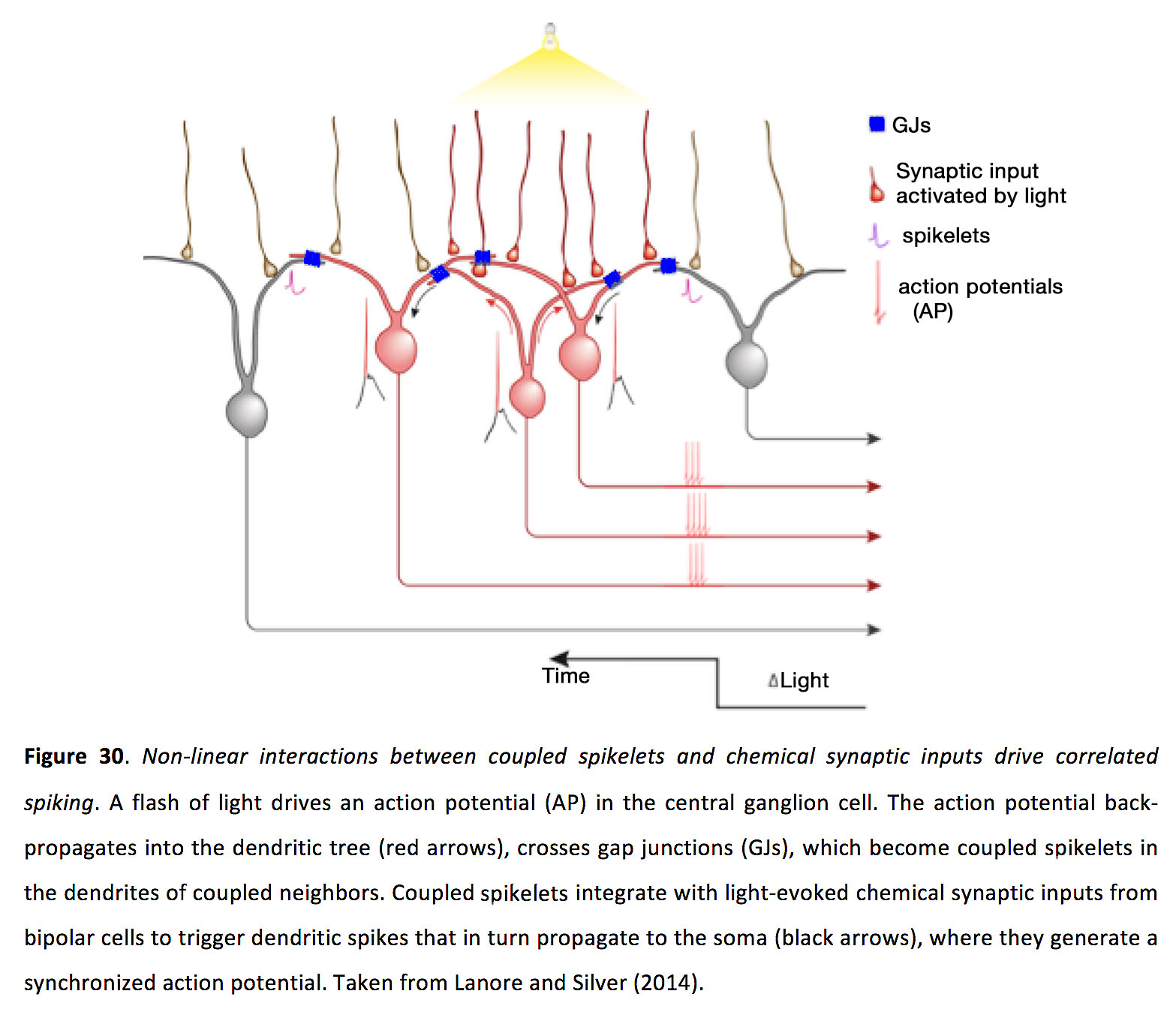 图30.耦合小穗和化学突触输入之间的非线性相互作用驱动相关穗.闪光驱动中枢神经节细胞的动作电位。动作电位反向传播到树突(红色箭头),穿过间隙连接(GJs),在耦合邻居的树突中成为耦合小穗。耦合的小穗与来自双极细胞的光诱发化学突触输入结合,触发树突小穗,进而传播到体细胞(黑色箭头),在那里它们产生同步的动作电位。取自Lanore和Silver(2014)(202)。
图30.耦合小穗和化学突触输入之间的非线性相互作用驱动相关穗.闪光驱动中枢神经节细胞的动作电位。动作电位反向传播到树突(红色箭头),穿过间隙连接(GJs),在耦合邻居的树突中成为耦合小穗。耦合的小穗与来自双极细胞的光诱发化学突触输入结合,触发树突小穗,进而传播到体细胞(黑色箭头),在那里它们产生同步的动作电位。取自Lanore和Silver(2014)(202)。
尽管它很流行,但相关的神经节细胞活动在高级视觉处理中扮演什么角色仍不清楚。一个假设的作用是,它可以增强视网膜受体细胞的突触传递。在侧膝状核(LGN)中,具有短峰间间隔的视网膜输入大大增加了突触的效能(151,152)。考虑到多个相邻的神经节细胞汇聚到单个LGN中继神经元(153-156),神经节细胞同步似乎是一个高度可信的策略,有效地驱动LGN神经元。神经节细胞同步的另一个假设作用是它提高视神经的带宽(145,157)。因为当光激活相邻神经节细胞感受野之间的重叠区域(图31)(145,157),相关活动包含独特的空间信息,随后可以与不相关活动分开解码。
 图31.相关的峰值携带着独特的空间信息.由闪烁的棋盘刺激产生的感受野被绘制为两个细胞(一个)和(C),而它们表现出相关峰值的“接受野”则显示在中间(B).D,接受野的高斯拟合轮廓(A-C)。顶部和底部的行来自不同的细胞对。摘自Meister et al(1995)(157)。
图31.相关的峰值携带着独特的空间信息.由闪烁的棋盘刺激产生的感受野被绘制为两个细胞(一个)和(C),而它们表现出相关峰值的“接受野”则显示在中间(B).D,接受野的高斯拟合轮廓(A-C)。顶部和底部的行来自不同的细胞对。摘自Meister et al(1995)(157)。
此外,由于窄间隙连接介导的神经节细胞相关性的强度似乎随着峰值率的增加而降低(143,158),尽管抑制机制有时可能同时降低峰值率并减少中等规模的间隙连接介导的同步性(159),相关的神经节细胞活性可以独立于峰值率向下游目标传递信息(图32).这些类型的并行解码策略是否真的在更高的视觉区域实现仍有待检验。
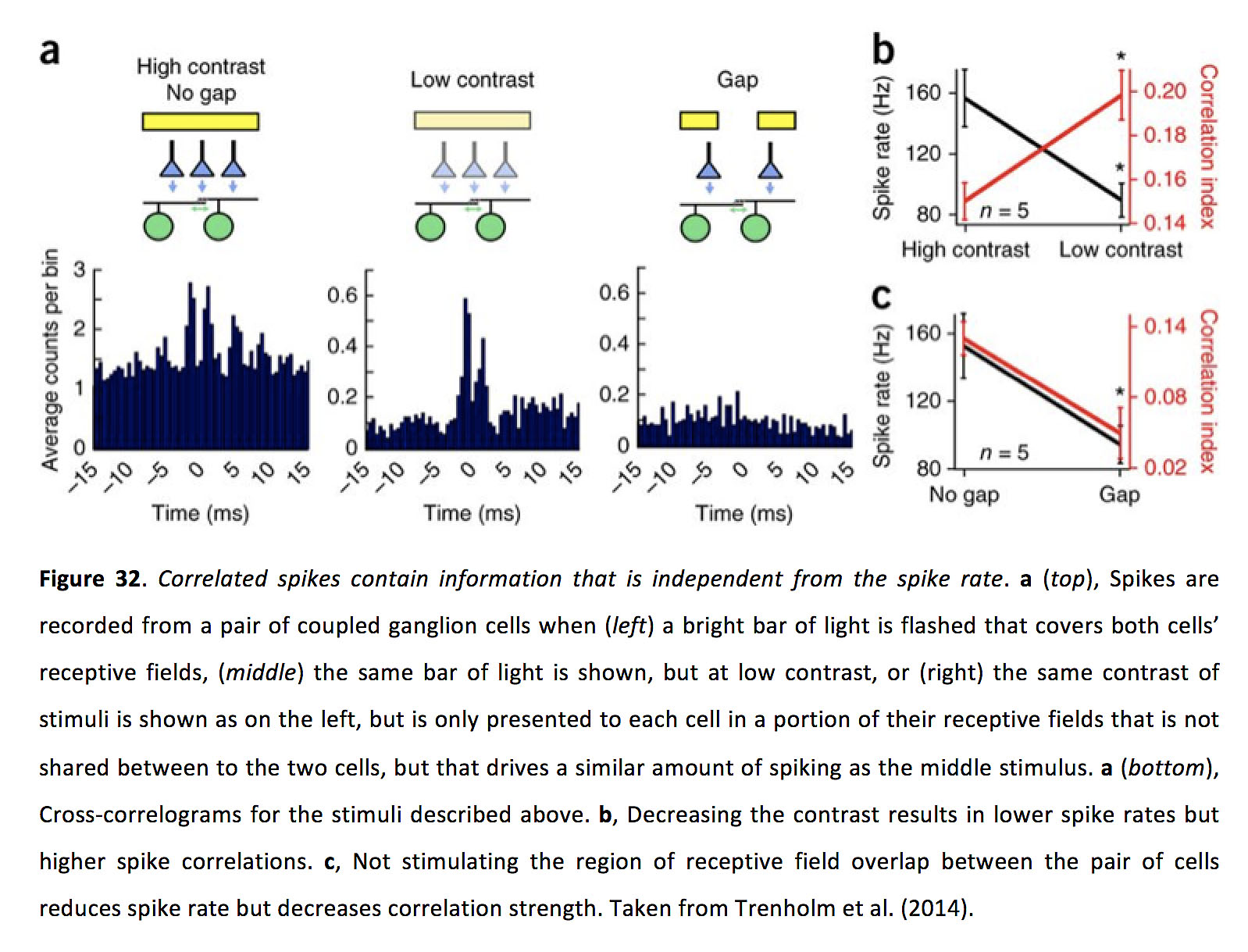 图32.相关的峰值包含独立于峰值速率的信息.一个(前),当(左)一道明亮的光柱照亮了两个细胞的感受区,(中间)显示相同的光条,但对比度较低,或(右)显示与左侧相同的刺激对比度,但只呈现给每个细胞的感受野的一部分,两个细胞之间不共享,但驱动的峰值量与中间刺激相似。一个(底)、上述刺激的交叉相关图。b,降低对比度会导致较低的峰值率,但较高的峰值相关性。c不刺激细胞对之间的感受野重叠区域可降低穗率,但降低相关强度。摘自Trenholm等人(2014)(143)。
图32.相关的峰值包含独立于峰值速率的信息.一个(前),当(左)一道明亮的光柱照亮了两个细胞的感受区,(中间)显示相同的光条,但对比度较低,或(右)显示与左侧相同的刺激对比度,但只呈现给每个细胞的感受野的一部分,两个细胞之间不共享,但驱动的峰值量与中间刺激相似。一个(底)、上述刺激的交叉相关图。b,降低对比度会导致较低的峰值率,但较高的峰值相关性。c不刺激细胞对之间的感受野重叠区域可降低穗率,但降低相关强度。摘自Trenholm等人(2014)(143)。
除了以上描述的同步形式(往往局限于邻近的视网膜神经元)外,间隙连接在视网膜大区域的同步活动中似乎也发挥着重要作用。配对记录的经典工作表明,视网膜上存在超过20度视觉空间(160度)的远程相关活动。当一对位于远端的神经节细胞对落在它们的感受野以及它们之间的空间上的视觉图像做出反应时,这种同步就出现了(160)。最近的研究表明,这种远程同步来自于神经节细胞之间的间隙连接连接,可能通过一个中间广域无腺细胞(161)。值得注意的是,正如最初的假设(160),这种远程同步似乎在动物将同一视觉图像的远端部分结合在一起的能力中发挥了重要的行为作用(161)。
缝隙连接和移动视觉刺激
兴奋性视网膜信号通路的经典观点是光感受器到双极细胞到神经节细胞的垂直回路。在相邻细胞之间放置间隙连接,增加了这条垂直通路的横向兴奋连接。不出所料,在处理移动的视觉刺激时,增加这样一个横向兴奋通路使视网膜回路具有明显的优势。
运动灵敏度
双极细胞和神经节细胞的侧间隙连接连接,已被证明有选择性地修改和增强对移动刺激的反应。在小鼠视网膜中,研究表明双极细胞的间隙连接输入(可代表其兴奋性突触输入的> %)可赋予双极细胞超线性空间总和,这可选择性地增强对时空相关输入的处理(即向一致方向移动的刺激)。这导致视网膜神经节细胞对运动的反应增强(图33)(84)。灵长类动物视网膜的相关研究表明,双极细胞间隙连接耦合似乎有助于太阳神经节细胞的运动敏感性(162)。双极细胞中缝隙连接网络运动促进的基本原理似乎与后面节细胞中描述的更详细的原理相似(163)。
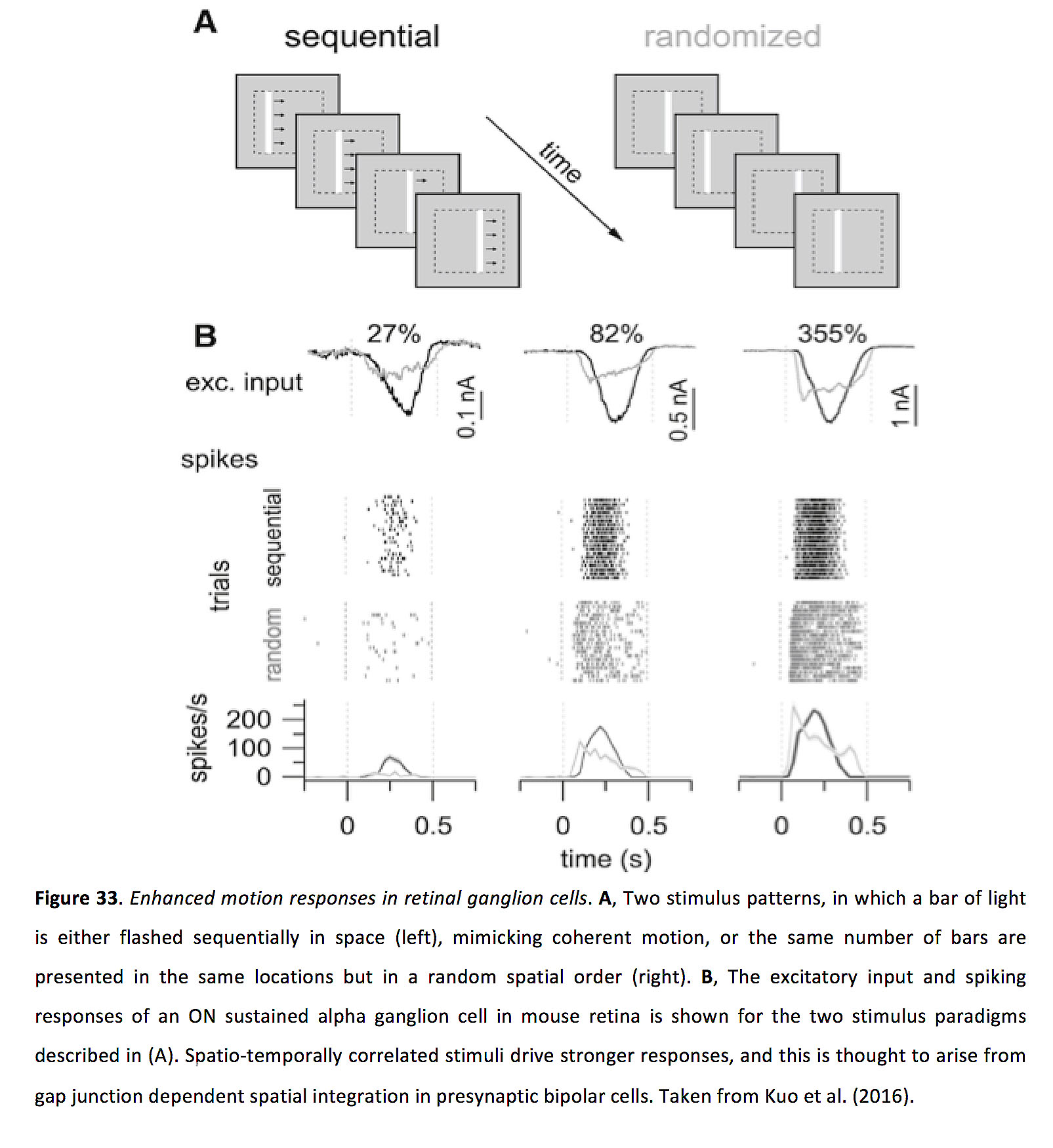 图33.增强视网膜神经节细胞的运动反应.一个,两种刺激模式,一束光在空间中连续闪烁(左),模仿连贯的运动,或者在相同的位置以随机的空间顺序呈现相同数量的光条(右)。B在(A)中描述的两种刺激范式中,小鼠视网膜上一个持续的α神经节细胞的兴奋性输入和峰值反应被显示出来。时空相关的刺激驱动更强的反应,这被认为来自于突触前双极细胞中间隙连接依赖的空间整合。摘自Kuo等人(2016)(84)。
图33.增强视网膜神经节细胞的运动反应.一个,两种刺激模式,一束光在空间中连续闪烁(左),模仿连贯的运动,或者在相同的位置以随机的空间顺序呈现相同数量的光条(右)。B在(A)中描述的两种刺激范式中,小鼠视网膜上一个持续的α神经节细胞的兴奋性输入和峰值反应被显示出来。时空相关的刺激驱动更强的反应,这被认为来自于突触前双极细胞中间隙连接依赖的空间整合。摘自Kuo等人(2016)(84)。
最后,在小鼠视网膜中,在低光照水平下,同源偶联方向选择神经节细胞比未偶联的神经节细胞具有更宽的调谐曲线。因此,缝隙连接连接似乎允许这些细胞在低光水平下,在探测微弱的移动刺激和准确报告移动方向之间取得平衡(图34)(111)。
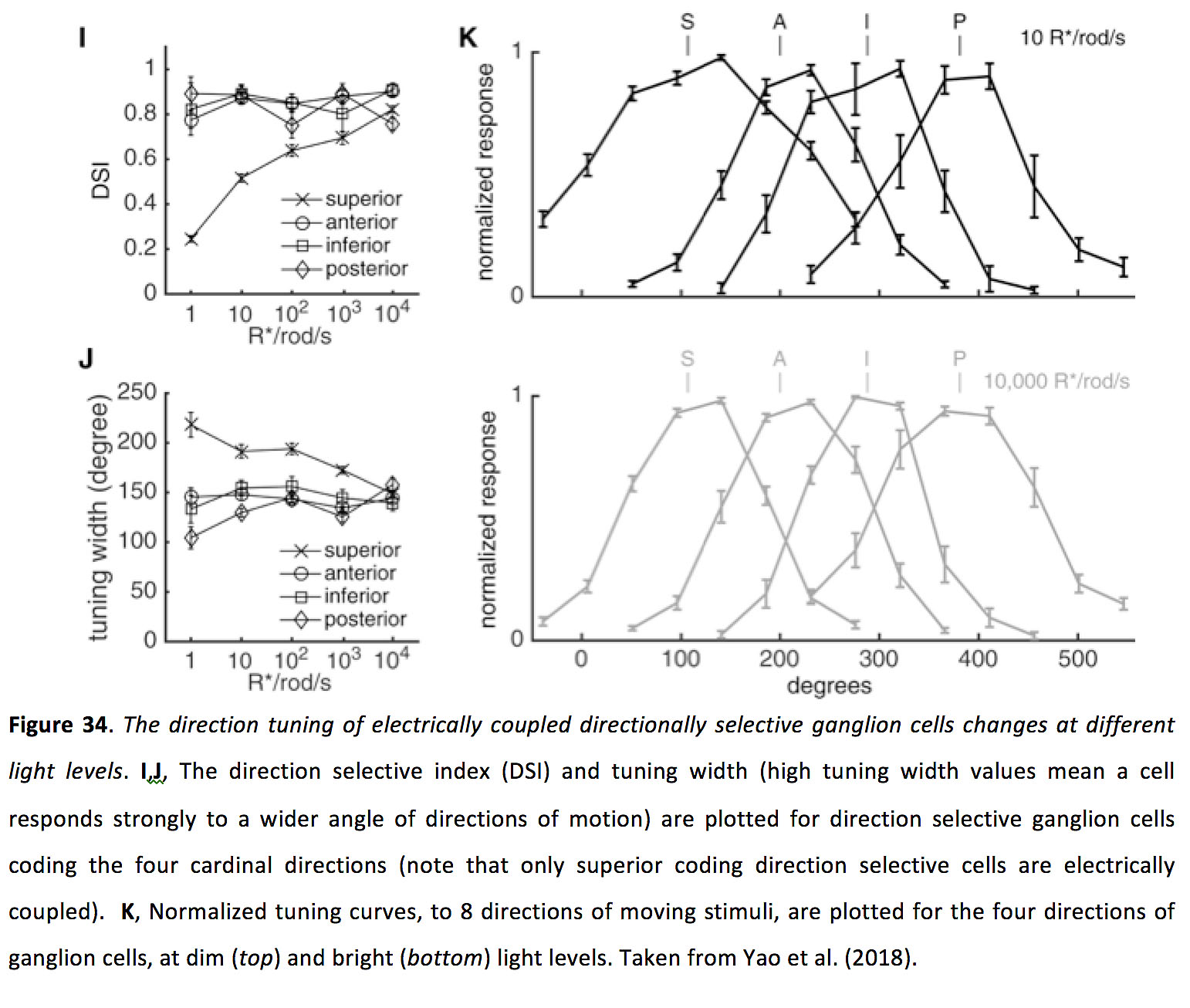
图34.电耦合定向选择神经节细胞的方向调谐在不同的光水平下发生变化.我,J,方向选择指数(DSI)和调节宽度(较高的调节宽度值意味着细胞对更大角度的运动方向有强烈的反应)被绘制为编码四个基本方向的方向选择神经节细胞(注意,只有较高级的编码方向选择细胞是电耦合的)。K神经节细胞在昏暗(前)和明亮的(底)光水平。摘自Yao等人(2018)(111)。
运动预期
当光在视网膜上闪现时,神经节细胞需要大约50毫秒或更长时间才能激发光诱发动作电位。这种延迟大部分是由于杆和锥的光转导,在这个相对缓慢的过程中,入射的光子被转换成电化学信号。这种延迟带来了一个问题,因为同一个视网膜环路可以处理静态和移动的刺激。对于静态图像来说,这种延迟并不会造成严重的问题,因为当神经节细胞的刺激信号达到峰值时,它所编码的刺激信号仍然处于适当的位置。相反,当一个移动的图像激活一个位置的感光细胞时,当垂直连接到较早位置的感光细胞的神经节细胞开始反应时,图像已经移动到不同的空间位置。随着运动速度的加快,神经节细胞报告图像的位置与图像在空间中的实际位置之间的时间差会增加。为了解决这一问题,视网膜已经假设了一种方法,即神经节细胞动态地将其感受野的峰值移向移动刺激的前沿(164,165),如果它们根据峰值峰值率解码位置,从理论上讲,下游目标可以读出这一峰值作为预期。另一种弥补编码移动刺激时产生的延迟的方法是利用缝隙连接,让上游的视网膜神经节细胞向下游的邻近细胞发出传入图像的警报。这种现象被称为滞后正常化,发生在同源偶联的视网膜神经节细胞中,这些细胞表现出偶联介导的阈下兴奋性感受野,包围着它们的感受野中心(图35)(113)。
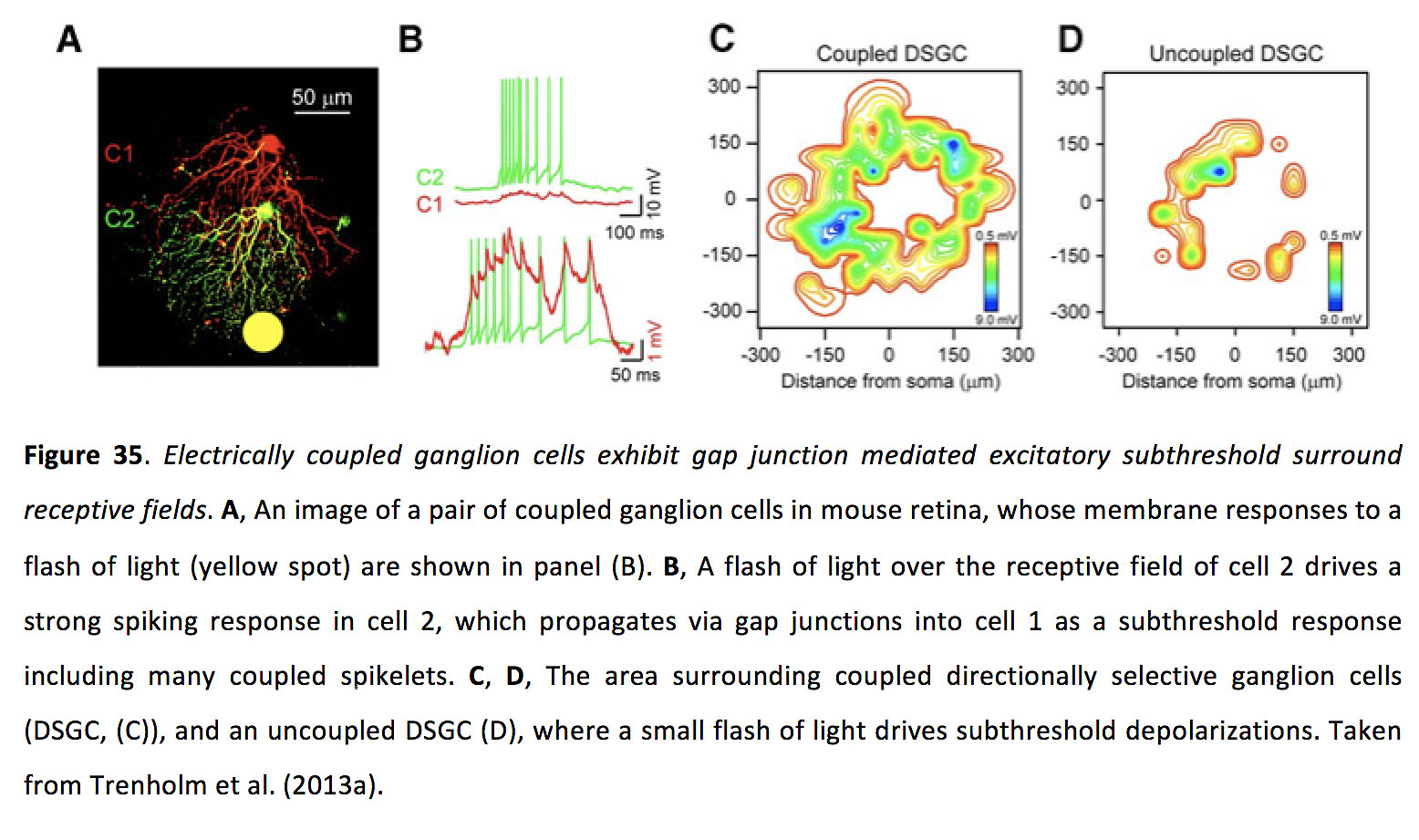 图35.电偶联神经节细胞表现为间隙连接介导的兴奋性阈下包围感受野.一个,小鼠视网膜上一对成对的神经节细胞的图像,其膜对闪光(黄色点)的反应如图(B)所示。B在细胞2的感受野上,一束光在细胞2的感受野上引发了强烈的峰值反应,并通过缝隙连接传播到细胞1,这是一种阈下反应,包括许多耦合的小穗。C,D,耦合定向选择性神经节细胞(DSGC, (C))周围的区域,和一个不耦合的DSGC (D),其中一个小闪光驱动阈下去极化。取自Trenholm等人(2013a)(113)。
图35.电偶联神经节细胞表现为间隙连接介导的兴奋性阈下包围感受野.一个,小鼠视网膜上一对成对的神经节细胞的图像,其膜对闪光(黄色点)的反应如图(B)所示。B在细胞2的感受野上,一束光在细胞2的感受野上引发了强烈的峰值反应,并通过缝隙连接传播到细胞1,这是一种阈下反应,包括许多耦合的小穗。C,D,耦合定向选择性神经节细胞(DSGC, (C))周围的区域,和一个不耦合的DSGC (D),其中一个小闪光驱动阈下去极化。取自Trenholm等人(2013a)(113)。
对于移动图像,间隙连接输入从一个神经节细胞横向扩散到另一个,并能够与弱双极细胞介导的化学突触输入相结合,使神经节细胞开始在接受区域外做出反应,从而驱动对静态刺激的峰值活动(166)。此外,当这些耦合的神经节细胞受到以不同速度移动的图像的刺激时,它们可以在几乎相同的空间位置启动反应(即“滞后规范化”),而非耦合的神经节细胞在看到速度越来越快的移动图像时,会在空间中越来越远的位置启动反应(图36)(166)。
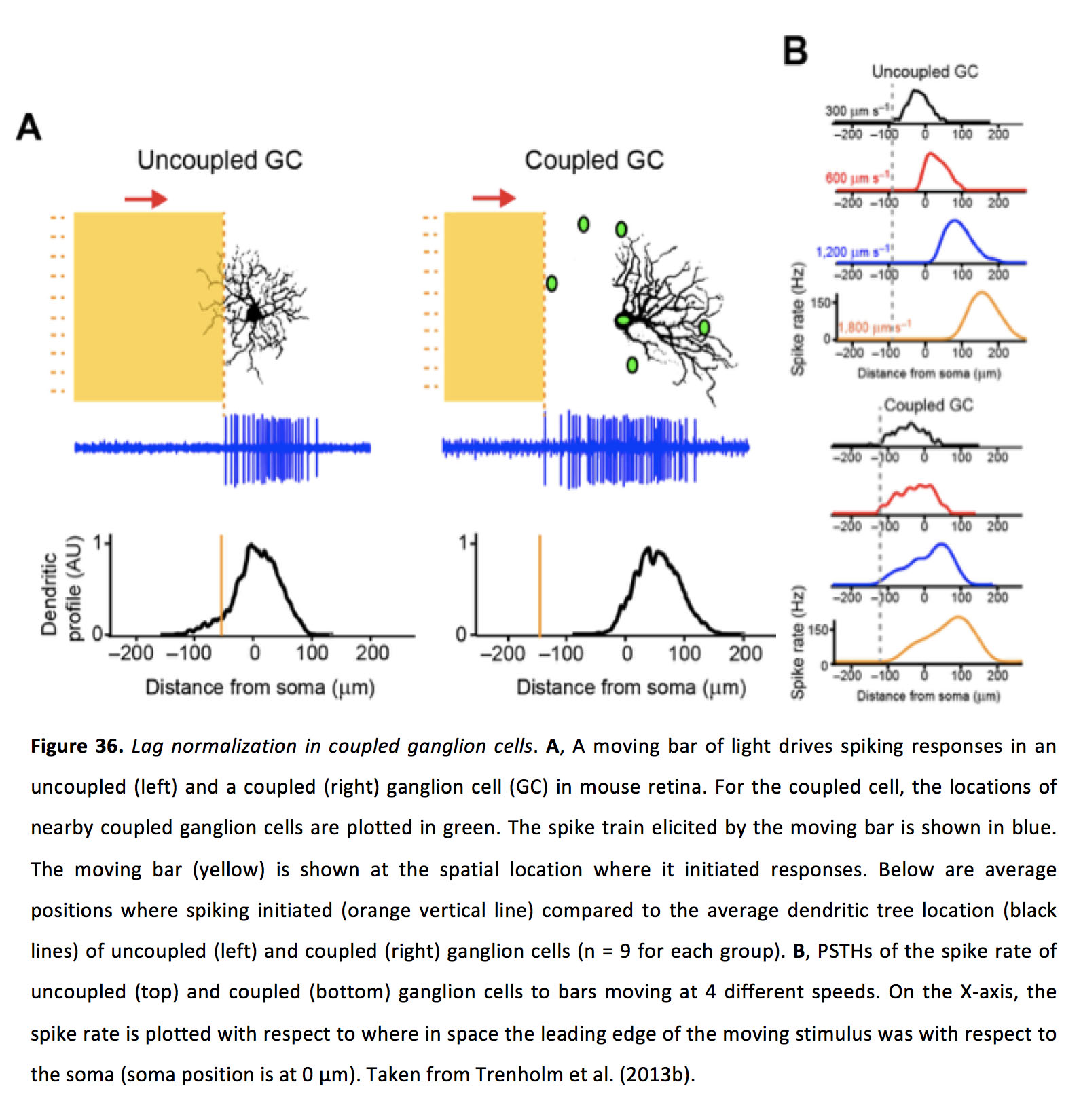 图36。偶联神经节细胞归一化滞后.一个在小鼠视网膜中,一个移动的光柱驱动未耦合(左)和耦合(右)神经节细胞(GC)的峰值反应。对于偶联细胞,附近的偶联神经节细胞的位置用绿色标出。由运动杆引起的尖刺序列显示为蓝色。移动条(黄色)显示在它发起响应的空间位置。下面是峰值开始的平均位置(橙色竖线)与未耦合(左)和耦合(右)神经节细胞的平均树突状树位置(黑色线)的对比(每组n = 9)。B,未偶联(上)和偶联(下)神经节细胞与以4种不同速度运动的棒状细胞的峰值速率的pths。在x轴上,脉冲速率与运动刺激的前缘在空间中的位置有关(体的位置为0 μ m)。摘自Trenholm等人(2013b)(166)。
图36。偶联神经节细胞归一化滞后.一个在小鼠视网膜中,一个移动的光柱驱动未耦合(左)和耦合(右)神经节细胞(GC)的峰值反应。对于偶联细胞,附近的偶联神经节细胞的位置用绿色标出。由运动杆引起的尖刺序列显示为蓝色。移动条(黄色)显示在它发起响应的空间位置。下面是峰值开始的平均位置(橙色竖线)与未耦合(左)和耦合(右)神经节细胞的平均树突状树位置(黑色线)的对比(每组n = 9)。B,未偶联(上)和偶联(下)神经节细胞与以4种不同速度运动的棒状细胞的峰值速率的pths。在x轴上,脉冲速率与运动刺激的前缘在空间中的位置有关(体的位置为0 μ m)。摘自Trenholm等人(2013b)(166)。
迫在眉睫的
有效地探测若隐若现的视觉刺激,比如逼近的捕食者,可能是生死攸关的问题。因此,若隐若现(或接近)的视觉刺激驱动了许多物种的行为反射,包括人类(167-171)。在视网膜中,所有的无分泌细胞和ON锥体双极细胞之间的侧隙连接似乎在使特定的视网膜神经节细胞优先检测若隐若离的视觉图像方面发挥了作用。在小鼠视网膜中,隐现的敏感细胞是OFF神经节细胞,称为PV-5细胞(124)。当这种细胞的感受野上出现静态的光衰减时,它们会引起短暂的OFF兴奋,然后当光水平恢复到基线(124)时,会引起短暂的ON抑制。当一个大的暗刺激在这个细胞的感受野上变小时,它只驱动ON抑制,而没有兴奋性的OFF反应,因此不会导致峰值。另一方面,如果同样的刺激现在向后播放,黑色区域扩展到细胞的感受野,它只驱动OFF激励,驱动健壮的峰值反应。根据所描述的反应特性,该细胞也应该对穿过感受野的移动刺激做出合理良好的反应,这将驱动OFF兴奋和on抑制的混合。然而,对PV-5细胞的ON抑制输入非常快,似乎是通过间隙连接信号从ON锥双极细胞传递到所有无分泌细胞而产生的,这削弱了伴随的兴奋性OFF输入的作用。事实上,当Cx36被敲除时,PV-5细胞不再优先对隐现的刺激作出反应,而对仅仅穿过感受野的刺激作出反应(124)。 In mice, this retinal cell type is thought to underlie innate freezing/fleeing behavioral responses when dark looming stimuli are presented above the head of a mouse (172). Whether similar retinal circuitry underlies behavioral responses to looming visual stimuli in primates remains to be tested.
缝隙连接,视网膜变性和自发性活动
虽然如上所述,间隙连接在健康视网膜中有许多功能,但它们也促进视网膜退行性变(rd)后的异常神经元活动。在啮齿类动物模型中,随着光感受器变性,许多神经节细胞不是沉默,而是自发振荡,频率约为10赫兹(图37)(173 - 179)。理解这种异常活动的来源很重要,因为它可能是视力丧失患者所经历的磷光烯的基础(180,181),并可能影响包括电植入(182)和光遗传视觉治疗(183)在内的治疗策略的成功。
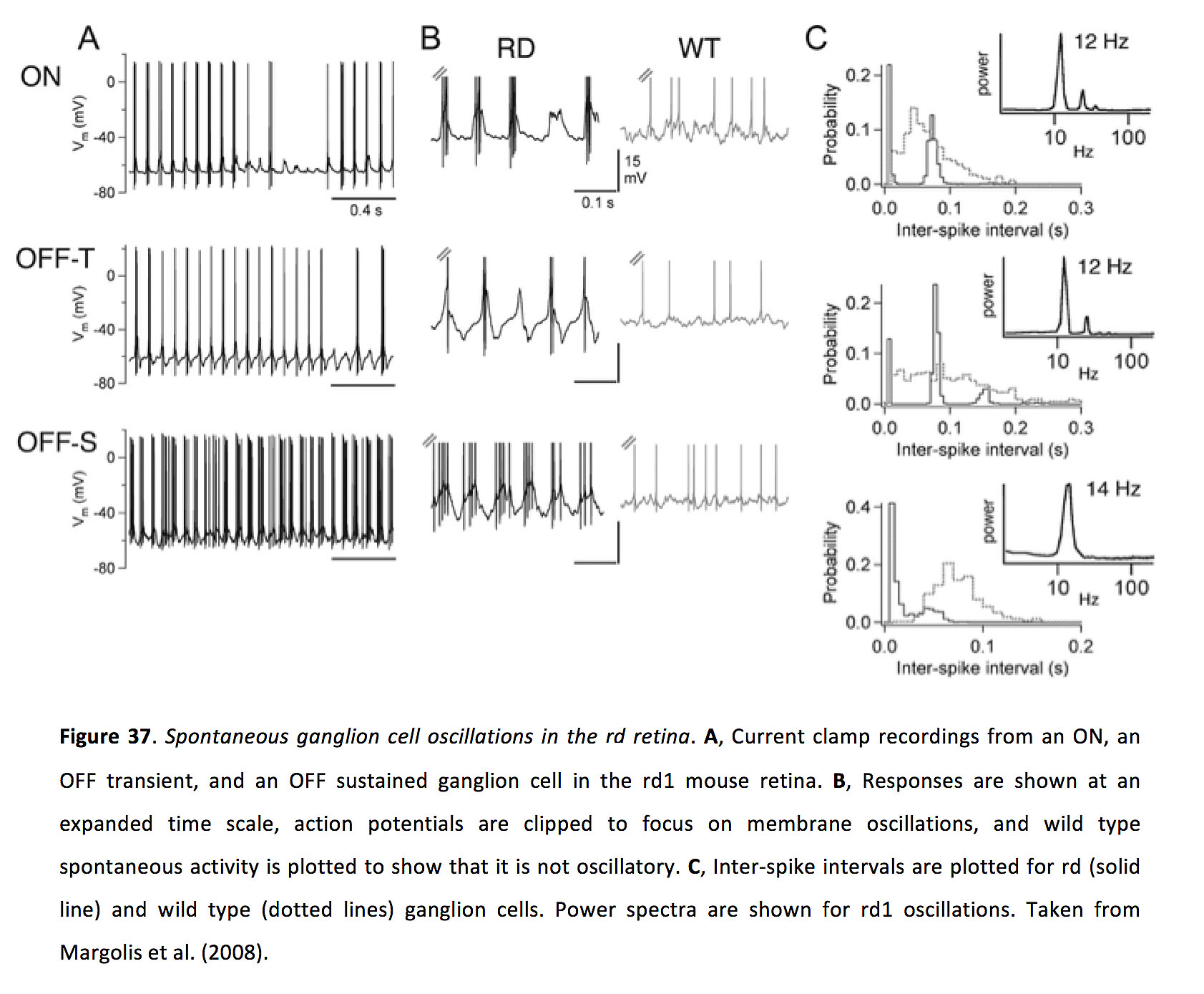 图37.视网膜视网膜内自发神经节细胞振荡.一个电流钳记录了rd1小鼠视网膜上的ON、OFF瞬态和OFF持续神经节细胞。B,反应在扩展的时间尺度上显示,动作电位被剪切以聚焦于膜振荡,野生型自发活动被绘制以表明它不是振荡的。Crd(实线)和野生型(虚线)神经节细胞的峰间间隔。给出了rd1振荡的功率谱。摘自Margolis等人(2008)(175)。
图37.视网膜视网膜内自发神经节细胞振荡.一个电流钳记录了rd1小鼠视网膜上的ON、OFF瞬态和OFF持续神经节细胞。B,反应在扩展的时间尺度上显示,动作电位被剪切以聚焦于膜振荡,野生型自发活动被绘制以表明它不是振荡的。Crd(实线)和野生型(虚线)神经节细胞的峰间间隔。给出了rd1振荡的功率谱。摘自Margolis等人(2008)(175)。
在视网膜中,神经节细胞由于接受兴奋性和抑制性振荡输入而振荡(174,175,177,184)。化学突触阻滞剂的应用抑制神经节细胞振荡,表明自发振荡并不发生在神经节细胞本身(174,176),但也见(185)。相反,ON锥双极细胞和所有无分泌细胞的膜振荡也以10hz左右的频率发生,在化学突触阻滞剂的作用下持续存在,表明自发振荡在ON锥双极细胞和所有无分泌细胞的耦合网络内内在地产生(174)。作为双极细胞和所有无分泌细胞之间的间隙连接在这种病理生理学中很重要的证据,药理学上阻断间隙连接可以抑制神经节细胞(176,186)、所有无分泌细胞和ON锥体双极细胞中的振荡(图38)(186)。此外,连接蛋白36被敲除后,rd小鼠自发的神经节细胞振荡大大减少(187)。
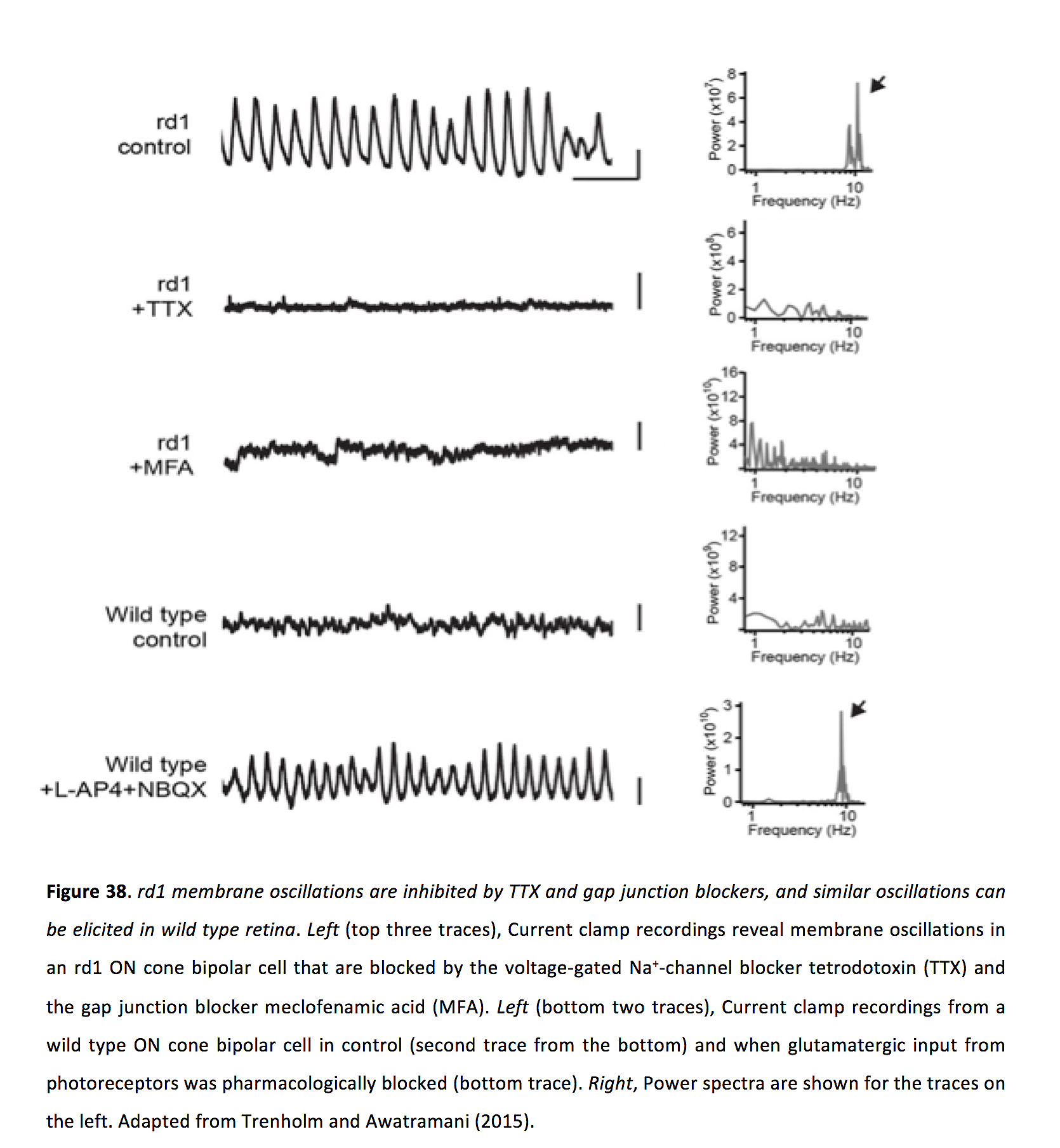 图38.rd1膜振荡被TTX和缝隙连接阻滞剂抑制,在野生型视网膜中也可诱发类似振荡。左(上面三个轨迹),电流钳记录显示rd1 ON锥双极电池中的膜振荡,被电压门控Na阻断+-通道阻滞剂河豚毒素(TTX)和缝隙连接阻滞剂甲氯胺酸(MFA)。左图(底部两个痕迹),电流钳记录的野生型ON锥双极细胞(底部第二个痕迹)和来自光感受器的谷氨酸输入被药理阻断时(底部的痕迹)。右,功率谱显示的痕迹在左边。改编自Trenholm和Awatramani(2015)(203)。
图38.rd1膜振荡被TTX和缝隙连接阻滞剂抑制,在野生型视网膜中也可诱发类似振荡。左(上面三个轨迹),电流钳记录显示rd1 ON锥双极电池中的膜振荡,被电压门控Na阻断+-通道阻滞剂河豚毒素(TTX)和缝隙连接阻滞剂甲氯胺酸(MFA)。左图(底部两个痕迹),电流钳记录的野生型ON锥双极细胞(底部第二个痕迹)和来自光感受器的谷氨酸输入被药理阻断时(底部的痕迹)。右,功率谱显示的痕迹在左边。改编自Trenholm和Awatramani(2015)(203)。
考虑到野生型视网膜中ON锥双极细胞和AII无腺细胞之间存在广泛的耦合,在视网膜变性过程中发生了什么,从而促进了异常的网络活动?野生型视网膜的实验表明,自发活动的主要触发因素与光感受器释放谷氨酸的改变有关,因为无论是药物阻断野生型视网膜的光感受器输出,还是用强光使野生型视网膜光脱色(两者都会使aiamacrine /ON锥体双极细胞网络超极化),都可以驱动自发活动,类似于rd视网膜中发现的自发活动(186)。尽管最近的研究也暗示维甲酸与这种过度活跃有关(188)。与前一观点一致的是,与野生型视网膜相比,变性视网膜中AII无分泌细胞的静息膜电位表现为超极化(189)。这种超极化可以激活节律性电导,比如Ih在一些ON锥双极细胞中存在,并已被证明可以调节rd视网膜的振荡(186)。这些振荡也被证明依赖于ttx敏感的Na+通道在所有细胞(186)。最后,研究表明,视网膜内所有无分泌细胞的间隙连接偶联似乎都增加了(通过磷酸化Cx36的增加进行测量),这可能加剧光感受器变性后的异常活性(190)。因此,AII双极细胞网络和ON双极细胞网络中的膜电导和间隙连接在光感受器变性后共同作用,引起自发的膜振荡(图39).
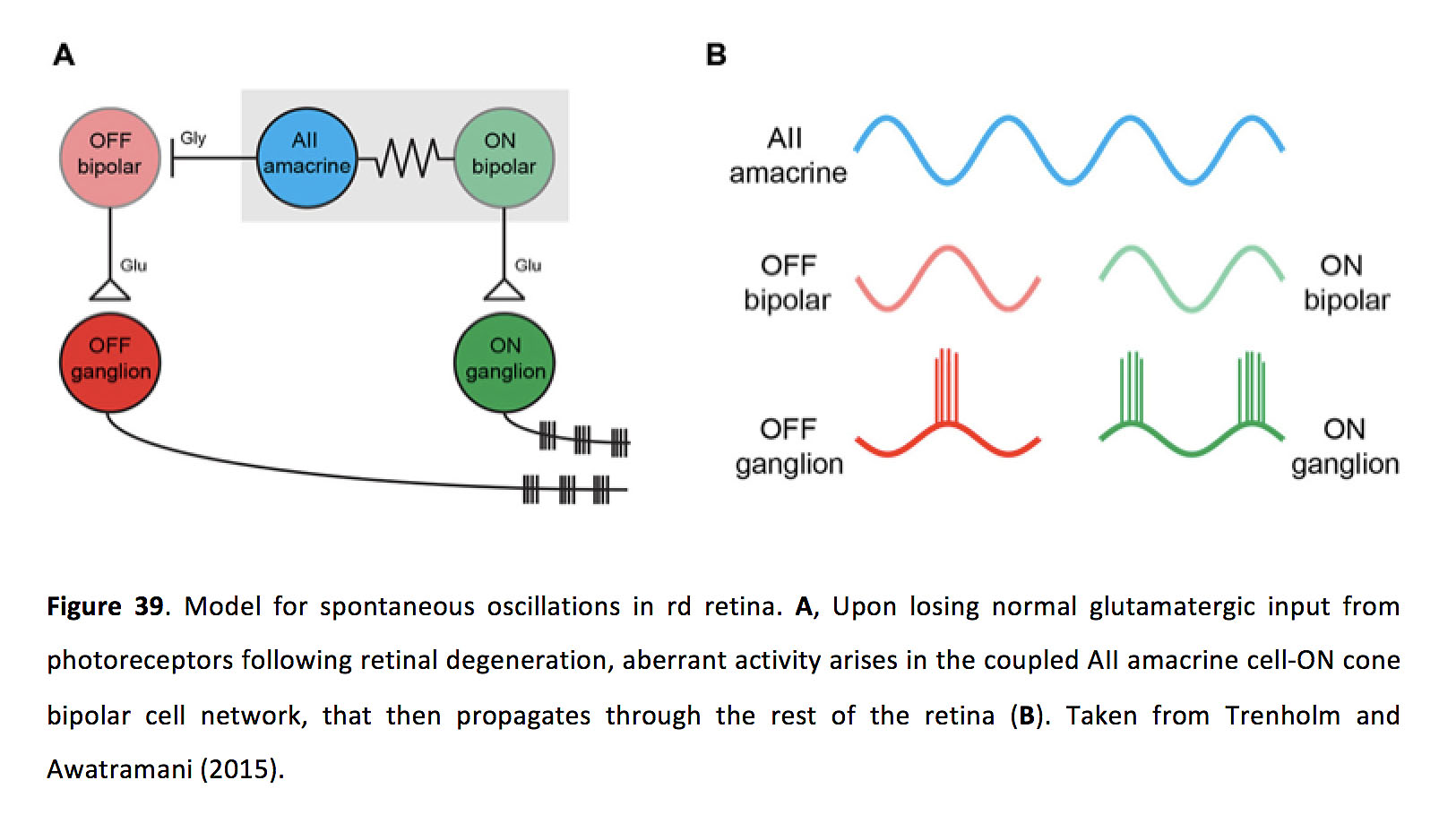 图39.视网膜自发振荡模型。一个视网膜变性后,当感光细胞失去正常的谷氨酸输入后,耦合的AII无分泌细胞-锥体双极细胞网络出现异常活动,然后通过视网膜的其余部分(B).取自Trenholm和Awatramani(2015)(203)。
图39.视网膜自发振荡模型。一个视网膜变性后,当感光细胞失去正常的谷氨酸输入后,耦合的AII无分泌细胞-锥体双极细胞网络出现异常活动,然后通过视网膜的其余部分(B).取自Trenholm和Awatramani(2015)(203)。
结论
已经确定的是,视网膜是广泛的间隙连接耦合,这些连接赋予视网膜电路大量的功能特性。在视网膜中,缝隙连接与夜视有关,控制感受野的大小、信号相关性、空间整合和运动检测。尽管有这些发现,仍有许多未知之处。首先,许多关于视网膜间隙连接作用的关键实验都使用了药理学制剂或cx36ko来检查视网膜回路中间隙连接的作用。然而,同时阻断视网膜所有层面的缝隙连接,使得在特定细胞类型之间分配特定电突触的作用变得困难。因此,未来的研究将需要努力修改特定细胞类型的缝隙连接(111)。第二,与第一点相关的是,虽然很明显,视网膜将传入的视觉信号分成30多个不同的视网膜通道(由不同的细胞类型组成,以不同的方式连接),但这些通道中缝隙连接的独特作用才刚刚开始研究。第三,鉴于间隙连接已知在视网膜发育中发挥重要作用(191),并可能影响疾病病理的易感性(192-195),研究间隙连接是否在发育和疾病中发挥精确的细胞类型特异性作用(196)将是重要的。最后,除了增加我们对缝隙连接在微电路中作用的细节知识外,重要的是要描述缝隙连接具有可塑性的不同方式,从而允许昼夜节律、光适应或神经元活动调节它们的活动并修改电路功能。
确认
这项工作得到了ST和GBA加拿大研究主席的支持。
关于作者

斯图尔特博士Trenholm在加拿大哈利法克斯的达尔豪西大学获得了神经科学博士学位,在高塔姆·阿瓦拉玛尼博士的实验室。随后,他在瑞士巴塞尔Friedrich Miescher生物医学研究所boond Roska博士的实验室做神经科学博士后。他现在是加拿大蒙特利尔麦吉尔大学蒙特利尔神经学研究所的助理教授。他的实验室检查了视觉知觉的神经回路,并研究了视觉康复策略。

博士Gautam Awatramani1995年获得罗切斯特大学理学士学位然后在纽约州立大学布法罗分校的马尔科姆·斯劳特博士的指导下获得了生理学和生物物理学博士学位。他在Vollum研究所(波特兰,2000-2004)与Laurence Trussell博士一起做博士后,在不列颠哥伦比亚大学(温哥华,bc省,2005-2007)与Tim Murphy博士一起做博士后,在Friedrich Miescher研究所(巴塞尔,CH, 2008)与Botond Roska博士一起做博士后。随后,他在达尔豪斯大学(哈利法克斯2008-2011)建立了自己的实验室,随后转到维多利亚大学(维多利亚,卑斯省),在那里他目前是生理学副教授/CRC主席。他目前的工作重点是理解视网膜产生方向选择性的突触机制。
参考文献
- Masland r.h.合著,视网膜的基本结构。自然神经科学杂志。2001;4(9): 877 - 886。(PubMed)
- Gollisch, T. M. Meister,比科学家认为的更聪明的眼睛:视网膜电路中的神经计算。神经元。2010;65(2): 150 - 164。(PubMed)
- Azeredo da Silveira, R.和B. Roska,电池类型,电路,计算。神经生物学最新观点。2011;21(5): 664 - 671。(PubMed)
- Masland r.h.合著,视网膜的神经元组织神经元。2012;76(2): 266 - 280。(PubMed)
- 钻石,刘昌明,视网膜中的抑制性中间神经元:类型、电路和功能。视觉科学年鉴。2017;3:1-24。(PubMed)
- 马特施特克、e.m.、广川K.E.、埃瓦茨、伯纳德、段x、李颖、吴良、吴世伟、B. Ouellette、J.J.罗亚尔、M. Stoecklin、王琼、曾浩、J.R. Sanes和J.A.哈里斯,视网膜神经节细胞的不同中央投射模式。细胞报告。2017;18日(8):2058 - 2072。(PubMed)
- 罗伯斯,E, E,劳雷尔和h,拜尔,视网膜投影组揭示了由神经节细胞多样性产生的大脑区域特异性视觉表征。当代生物学:CB。2014;24(18): 2085 - 2096。(PubMed)
- Ribelayga, C.P.和J. O 'Brien,第十章-脊椎动物视网膜电突触可塑性的昼夜节律和光适应控制,在网络功能与可塑性荆洁,编者。2017,学术出版社。p . 209 - 241。(PubMed)
- 贝勒,地方检察官,m·g·福尔特斯,还有奥布赖恩,海龟视网膜上的视锥细胞接受区。《生理学杂志》1971;214(2): 265 - 294。(PubMed)
- 哥本哈根,D.R.和W.G.欧文,脊椎动物视网膜中杆状感光器之间的耦合大自然。1976;260(5546): 57-59。(PubMed)
- 卡斯特,n.v.蝾螈视网膜感光细胞之间结构上的特殊接触。比较神经学杂志1973;151(1): 35-56。(PubMed)
- 德弗里斯,s.h., X.祁,R.史密斯,W.马库斯,P.斯特林,哺乳动物锥体之间的电耦合。当代生物学:CB。2002;12(22): 1900 - 1907。(PubMed)
- 霍恩斯坦,e.p., J. Verweij, P.H. Li和J. l . Schnapf,猕猴视网膜间隙连接耦合和光感受器的绝对敏感性。神经科学杂志:神经科学学会官方杂志,2005;25(48): 11201 - 11209。(PubMed)
- 霍恩斯坦,e.p., J. Verweij和J. l . Schnapf,灵长类动物视网膜中红色和绿色视锥细胞之间的电耦合。自然神经科学杂志。2004;7(7): 745 - 750。(PubMed)
- 李,p.h., J. Verweij, J. h . Long和J. l . Schnapf,哺乳动物杆状光感受器的间隙-连接耦合及其对视觉检测的影响。《神经科学学报》(英文版)2012;32(10): 3552 - 3562。(PubMed)
- Raviola, E.和nb . Gilula,脊椎动物视网膜中感光细胞之间的缝隙连接。《美国国家科学院学报》1973;70(6): 1677 - 1681。(PubMed)
- 冢本,Y., K. Morigiwa, M. Ueda, P. Sterling小鼠视网膜夜视微电路。《神经科学杂志:美国神经科学学会官方杂志》2001;21日(21):8616 - 8623。(PubMed)
- 费根span, U. Janssen-Bienhold, S. Hormuzdi, H. Monyer, J. Degen, G. Söhl, K. Willecke, J. Ammermüller, R. Weiler,小鼠视网膜锥蒂和离锥双极细胞中连接蛋白36的表达。《神经科学杂志:美国神经科学学会官方杂志》2004;24(13): 3325 - 3334。(PubMed)
- Kántor, O, Z.本卡托斯,A. Énzsöly, C. Dávid, A.瑙曼,R.尼奇克,A. Szabó, E. Pálfi, J. Orbán, M.尼特拉伊,J. Németh, Á。Szel,。Lukáts和B. Völgyi,人外视网膜中连接隙连接的特征。脑结构与功能。2016;221(6): 2963 - 2984。(PubMed)
- 李,E.-J。, J.-W。汉族,周宏儒。金,I.-B。金,M.-Y。李,S.-J。哦,J.-W。钟,M.-H。春,连接蛋白36在豚鼠视网膜杆和锥间隙连接处的免疫细胞化学定位。欧洲神经科学杂志,2003;18(11): 2925 - 2934。(PubMed)
- 奥布莱恩,j.j., x.c Chen, P.R.麦克利什,J.奥布莱恩,S.C.梅西,灵长类视网膜中由连接蛋白36介导的光感受器耦合。《神经科学学报》(英文版)2012;32(13): 4675 - 4687。(PubMed)
- 李浩,庄亚正,j.o 'Brien,斑马鱼视网膜上的光感受器偶联受连接蛋白35磷酸化控制。神经科学杂志:神经科学学会官方杂志。2009;29(48): 15178 - 15186。(PubMed)
- Li, H, Zhang z, mr . r . Blackburn, s.w Wang, C.P. Ribelayga, J. O 'Brien,小鼠视网膜中腺苷和多巴胺受体通过缝隙连接磷酸化调节光感受器偶联。神经科学学报:美国神经科学学会官方杂志。2013;33(7): 3135 - 3150。(PubMed)
- 张俊,吴绍明,Connexin35/36间隙连接蛋白在虎蝾螈视网膜的光感受器中表达。比较神经学杂志。2004;470(1): 1 - 12。(PubMed)
- S. Asteriti, C. Gargini和L. Cangiano,连接蛋白36的表达是小鼠杆和锥之间电耦合所必需的。视觉神经科学。2017;34: E006。(PubMed)
- 科尔布、H。猫视网膜外丛状层的组织:电子显微镜观察。神经细胞学杂志1977;6(2): 131 - 153。(PubMed)
- 李,W.和S.H.德弗里斯,哺乳动物视网膜上分开的蓝色和绿色锥体网络。自然神经科学杂志。2004;7(7): 751 - 756。(PubMed)
- 冢本,Y., P. Masarachia, S.J. Schein和P. Sterling,猕猴中央凹锥花梗之间的间隙连接。视觉研究。1992;32(10): 1809 - 1815。(PubMed)
- 张俊,吴绍明,虎蝾螈视网膜杆状光感受器电偶联的生理特性。生理学杂志,2005;564 (Pt 3): 849 - 862。(PubMed)
- Werblin f.s.,在虎蝾螈视网膜中沿着杆状细胞和杆状细胞之间传播。生理学杂志。1978;280:449 - 470。(PubMed)
- 高,F。,j j。和吴少明,暗适应视网膜中杆状光感受器和锥状光感受器之间的信号保留和信号反转突触相互作用。生理学报,2013;591(22): 5711 - 5726。(PubMed)
- Attwell, D., M. Wilson, and S.M. Wu,蝾螈视网膜光感受器相互作用的定量分析。《生理学杂志》1984;352:703 - 737。(PubMed)
- Ribelayga, C., Y. Cao和S.C. Mangel,视网膜中的生物钟控制杆锥耦合。神经元。2008;59(5): 790 - 801。(PubMed)
- 吴绍明、杨晓亮、蝾螈视网膜中视杆和视锥之间的电耦合。《美国国家科学院院刊》1988;85(1): 275 - 278。(PubMed)
- S. Asteriti, C. Gargini和L. Cangiano,老鼠的杆状细胞通过与视锥细胞的缝隙连接发出信号。eLife。2014;3: e01386。(PubMed)
- 纳尔逊,R。猫视锥细胞有杆状输入:猫视网膜视锥细胞和水平细胞体反应特性的比较。比较神经学杂志1977;172(1): 109 - 135。(PubMed)
- Schneeweis, D.M.和J.L. Schnapf,猕猴视网膜视杆和视锥细胞的光电压。科学(纽约,纽约州)。1995;268(5213): 1053 - 1056。(PubMed)
- Thoreson, W.B.和S.C. Mangel,视网膜外侧的横向相互作用。视网膜与眼科学研究进展。2012;31(5): 407 - 441。(PubMed)
- Raviola, E.和nb . Gilula,视网膜外丛状层中特殊接触的膜内组织。猴子和兔子的冷冻骨折研究。《细胞生物学杂志》,1975;65(1): 192 - 222。(PubMed)
- 霍巴赫、詹森-比恩霍尔德、G. Söhl、T.舒伯特、H. Büssow、T.奥特、R.维勒和K.威勒克,连接蛋白57在小鼠视网膜水平细胞中的功能表达。《欧洲神经科学杂志》2004;19日(10):2633 - 2640。(PubMed)
- 杨森-比恩霍尔德,J. Trümpler, G.希尔根,K.舒尔茨,L.P.D.S. Müller, S.桑塔格,K.德德克,P.德克斯,K.威勒克,R.韦勒,Connexin57在小鼠水平细胞的树突-树突和轴突-轴突间隙连接中表达,其分布受光调节。比较神经病学杂志。2009;513(4): 363 - 374。(PubMed)
- Dorgau, B., R. Herrling, K. Schultz, H. Greb, J. Segelken, S. Ströh, P. Bolte, R. Weiler, K. Dedek,和U. Janssen-Bienhold,小鼠水平细胞轴突末端通过同型间隙连接连接50对偶联。比较神经病学杂志。2015;523(14): 2062 - 2081。(PubMed)
- O 'Brien, j.j., Li W., Pan F., J. Keung, J. O 'Brien, S.C. Massey,在兔视网膜中,a型水平细胞之间的耦合是由连接蛋白50间隙连接介导的。《神经科学杂志:神经科学学会官方杂志》2006;26日(45):11624 - 11636。(PubMed)
- 潘芳芳,姜志强,李毅。金,M.B.库斯,S.L.米尔斯,J.奥布莱恩和S.C.梅西,连接蛋白57通过兔视网膜b型水平细胞轴突末端网络表达。比较神经病学杂志。2012;520(10): 2256 - 2274。(PubMed)
- 克拉森、l.j.、孙卓、M.N.施泰雅特、P.博尔特、i.f仑福特、T. Sjoerdsma、J. Klooster、Y. Claassen、C.R. Shields、H.M.M. Ten Eikelder、U. Janssen-Bienhold、G. Zoidl、D.G. McMahon和M. Kamermans,由于连接蛋白半通道的缺失,从水平细胞到锥细胞的突触传递受到损害。公共科学图书馆生物学》杂志上。2011;9 (7): e1001107。(PubMed)
- Shields, C.R, J. Klooster, Y. Claassen, M. Ul-Hussain, G. Zoidl, R. Dermietzel和M. Kamermans,斑马鱼连接蛋白52.6和55.5视网膜水平细胞特异性启动子活性及蛋白表达。《比较神经病学杂志》2007;501(5): 765 - 779。(PubMed)
- 德弗里斯,S.H.和E.A.施瓦茨,多巴胺和第二信使对鲶鱼水平细胞间电突触的调节。生理学杂志,1989;414:351 - 375。(PubMed)
- Lasater, E.M.和J.E. Dowling,多巴胺降低培养的视网膜水平细胞间电连接的电导。《美利坚合众国国家科学院院刊》,1985;82(9): 3025 - 3029。(PubMed)
- 麦克马洪,D.G斑马鱼视网膜水平细胞电突触传递的调节。神经科学杂志:神经科学学会官方杂志。1994;14 (3 Pt 2): 1722 - 1734。(PubMed)
- 麦克马洪,D.G.和M.P.马特森,巨丹尼奥的水平细胞电耦合:由多巴胺调节突触和由钙维持突触。大脑研究。1996;718(2): 89 - 96。(PubMed)
- Xin, D.和S.A. Bloomfield,哺乳动物视网膜水平细胞间由暗、光引起的耦合变化。《比较神经病学杂志》1999;405(1): 75 - 87。(PubMed)
- Ribelayga和S.C. Mangel,水平细胞间隙连接耦合的生物钟调节缺失揭示了金鱼视网膜中的两个多巴胺系统。比较神经病学杂志。2003;467(2): 243 - 253。(PubMed)
- Ribelayga和S.C. Mangel,鱼竿水平细胞之间的示踪耦合:由光和多巴胺调制,但不受视网膜昼夜时钟的影响。视觉神经科学。2007;24(3): 333 - 344。(PubMed)
- 曼格尔,S.C.和J.E.道林,鲤鱼水平细胞的反应性和感受野大小被延长的黑暗和多巴胺减少。科学(纽约,纽约州)。1985;229(4718): 1107 - 1109。(PubMed)
- 雪莱、J、K.德德克、T.舒伯特、A.费根斯潘、K.舒尔茨、S.霍姆巴赫、K.威勒克和R.维勒,连接蛋白57缺陷小鼠水平细胞感受野减少。《欧洲神经科学杂志》2006;23日(12):3176 - 3186。(PubMed)
- 甜菜,南卡罗来纳州。兔视网膜神经节细胞周围感受野水平细胞贡献的分析。生理学杂志。1991;442(1): 211 - 234。(PubMed)
- 沃布林,F.S.和J.E.道林,泥鳅视网膜的组织,黄斑泥鳅。2细胞内记录。神经生理学杂志。1969;32(3): 339 - 355。(PubMed)
- 德里南伯格,F.弗兰克,R.K.森川,J. Jüttner, D.希利尔,P.汉茨,A.耶罗曼,R.阿泽雷多·达·西尔韦拉,B.罗斯卡,第一个视觉突触的反馈如何产生不同的视网膜功能。神经元。2018;99 (1): 117 - 134. - e11。(PubMed)
- 法罗,K., M.特谢拉,T.希克拉,T. j . Viney, K.巴林特,K. Yonehara和B. Roska,环境照明拨动视网膜中的神经元电路开关,使视觉感知达到锥体阈值。神经元。2013;78(2): 325 - 338。(PubMed)
- 霍加斯,A.J.麦克劳克林,K.罗内伦菲奇,S.特伦霍尔姆,R.瓦桑达尼,S.塞图拉马努贾姆,D.施瓦布,K. l .布里格曼,G.B.阿瓦拉马尼,在定向选择视网膜回路中,不同的无分泌细胞的特定线路允许独立的方向和大小编码。神经元。2015;86(1): 276 - 291。(PubMed)
- Dedek, K. Pandarinath, N.M. Alam, K. Wellershaus, T. Schubert, K. Willecke, G.T. Prusky, R. Weiler和S. Nirenberg,神经节细胞适应性:水平细胞的耦合是否起作用?《公共科学图书馆•综合》。2008;3 (3): e1714。(PubMed)
- 潘达里纳斯,C., I.波马什,J.D.维克多,G.T.普鲁士基,W.W.切特和S.尼伦伯格,一种将神经系统从一种状态切换到另一种状态的新机制。计算机科学进展。2010;2节。(PubMed)
- Wässle, H., C. Puller, F. Müller, S. Haverkamp,小鼠视网膜中双极细胞的锥接触、镶嵌和领地。神经科学杂志:神经科学学会官方杂志。2009;29(1): 106 - 117。(PubMed)
- Kolb H.和E.V. Famiglietti,猫视网膜内丛状层的杆状和锥状通路。科学(纽约,纽约州)。1974;186(4158): 47-49。(PubMed)
- 李,s.c.s.,梅耶,舒伯特,L. Hüser, K. Dedek和S. Haverkamp,小鼠视网膜A8双层小无分泌细胞的形态和连通性。比较神经病学杂志。2015;523(10): 1529 - 1547。(PubMed)
- 新井先生,田中先生,立花先生,电偶联双极细胞网络在成人视网膜中的主动作用。《神经科学杂志:美国神经科学学会官方杂志》2010;30(27): 9260 - 9270。(PubMed)
- 雅各比,R.A.和D.W.马尔沙克,猕猴视网膜DB3弥漫性双极细胞轴突的突触连接。比较神经病学杂志,2000;416(1): 19-29。(PubMed)
- Marc, r.e., W.L. Liu和J.F. Muller,金鱼视网膜内丛状层的缝隙连接。视觉研究。1988;28(1): 9-24。(PubMed)
- 斯特莱托,E, E, Raviola和r。f。达切,兔视网膜内窄场双层棒无分泌细胞(AII)的突触连接。比较神经病学杂志。1992;325(2): 152 - 168。(PubMed)
- 迪恩斯,m.r., B. Volgyi, D.A. Goodenough, s.a Bloomfield, D.L. Paul,在哺乳动物的视网膜中,连接蛋白36对于杆状细胞介导的视觉信号的传输至关重要。神经元。2002;36(4): 703 - 712。(PubMed)
- Han y和S.C. Massey,视网膜ON锥双极细胞中的电突触:连接蛋白的亚型特异性表达。美国国家科学院学报,2005;102(37): 13313 - 13318。(PubMed)
- Hilgen, G, J. von Maltzahn, K. Willecke, R. Weiler和K. Dedek,小鼠视网膜OFF双极细胞中连接蛋白45的亚细胞分布。比较神经病学杂志。2011;519(3): 433 - 450。(PubMed)
- 马克西纳,S., K.德德克,U.詹森-比恩霍尔德,J. Ammermüller, H.布伦,T.基尔施,M.皮珀,J.德根,O. Krüger, K.威勒克,R.维勒,小鼠视网膜神经元连接蛋白45的缺失破坏了双极细胞间的杆状/锥状信号通路,导致视觉传递受损。神经科学杂志:神经科学学会官方杂志,2005;25(3): 566 - 576。(PubMed)
- 达西,D., O.S.帕克,L.迪勒,D.布雷纳德,B.彼得森和B.李,灵长类动物视网膜锥体双极细胞的中心环绕感受野结构。视觉研究。2000;40(14): 1801 - 1811。(PubMed)
- Kujiraoka和T. Saito,鲤鱼视网膜双极细胞间的电偶联。《美利坚合众国国家科学院院刊》,1986;83(11): 4063 - 4066。(PubMed)
- 米尔斯,S.L。兔视网膜中锥状双极细胞的异常耦合模式。视觉神经科学。1999;16(6): 1029 - 1035。(PubMed)
- 明野,O.,前原M., S. Hidaka, S. Kita, Y.桥本,硬骨鱼视网膜双极-双极细胞偶联的网络特性。视觉神经科学。1994;11(3): 533 - 548。(PubMed)
- Veruki M.L.和E. Hartveit,在哺乳动物的视网膜中,大泡细胞形成一个电偶联的中间神经元网络。神经元。2002;33(6): 935 - 946。(PubMed)
- 马克,r.e., J.R.安德森,B.W.琼斯,C.L. Sigulinsky和J.S. Lauritzen,全无分泌细胞连接组:密集的网络集线器。神经网络学报,2014;8:104。(PubMed)
- 格雷顿,c.w., E.E.利伯曼,n·罗,k·l·布里格曼,j·h·辛格,j·s·戴蒙德,小鼠视网膜中全无分泌细胞介导的杆状通路和锥状通路之间的突触转移。当代生物学:CB。2018;28 (17): 2739 - 2751. - e3。(PubMed)
- Saito, T.和T. Kujiraoka,鲤鱼视网膜的双极-双极耦合特征。《普通生理学杂志》1988;91(2): 275 - 287。(PubMed)
- Berntson和W.R. Taylor,小鼠视网膜上双极细胞的反应特性和感受野宽度。生理学杂志,2000;524 Pt 3:879 - 889。(PubMed)
- Werblin f.s.,视网膜细胞对运动点的反应:黄斑Necturus细胞内记录。神经生理学杂志。1970;33(3): 342 - 350。(PubMed)
- 郭,S.P, G.W. Schwartz和F. Rieke,视网膜电和化学突触的非线性时空整合。神经元。2016;90(2): 320 - 332。(PubMed)
- Masland r.h.合著,无分泌细胞的任务。视觉神经科学。2012;29(1): 3 - 9。(PubMed)
- 马斯兰德,R.H.和J.W.米尔斯,兔视网膜中乙酰胆碱的放射自显影鉴别。细胞生物学杂志,1979;83(1): 159 - 178。(PubMed)
- Lee, S., K. Kim和Z.J. Zhou,ACh-GABA共传输在图像运动和运动方向检测中的作用。神经元。2010;68(6): 1159 - 1172。(PubMed)
- Sethuramanujam, S, A.J. McLaughlin, G. deRosenroll, A. Hoggarth, D.J. Schwab和G. b . watramani,乙酰胆碱/GABA混合传输在视网膜定向编码中的中心作用。神经元。2016;90(6): 1243 - 1256。(PubMed)
- 张,D.-Q。, T.-R。周和D.G.麦克马洪,视网膜多巴胺能神经元在视觉中的多种作用的功能异质性。《神经科学杂志:美国神经科学学会官方杂志》2007;27(3): 692 - 699。(PubMed)
- 巴登,t和t,欧拉,视网膜生理学:内视网膜的非双极细胞兴奋驱动。当代生物学:CB。2016;26日(15):R706-R708。(PubMed)
- Xin, D.和S.A. Bloomfield,兔视网膜无分泌细胞与神经节细胞的示踪偶联模式。比较神经病学杂志。1997;383(4): 512 - 528。(PubMed)
- Li, W, J. Zhang和S.C. Massey,兔视网膜S1和S2无汗腺细胞的耦合模式。视觉神经科学。2002;19(2): 119 - 131。(PubMed)
- 阿卜杜勒-马吉德,r.m., M.L.阿奇博尔德,F.特伦布莱,W.H.鲍德里奇,荧光金标记大鼠视网膜内核层神经元的示踪偶联。大脑研究。2005;1063(2): 114 - 120。(PubMed)
- 布卢姆菲尔德和D.辛,兔视网膜无腺细胞和神经节细胞受体和示踪耦合大小的比较。视觉神经科学。1997;14(6): 1153 - 1165。(PubMed)
- Völgyi, B., S. Chheda和S.A. Bloomfield,小鼠视网膜神经节细胞亚型的示踪偶联模式。比较神经病学杂志。2009;512(5): 664 - 687。(PubMed)
- 潘,F., D.L.保罗,S.A.布卢姆菲尔德和B. Völgyi,在小鼠视网膜中,大多数神经节细胞亚型的间隙连接耦合需要连接蛋白36。比较神经学杂志,2010;518(6): 911 - 927。(PubMed)
- 庞,j j。,D.L. Paul, and S.M. Wu,小鼠视网膜内无分泌细胞与逆行性神经节细胞偶联的研究。眼科与视觉科学,2013;54(8): 5151 - 5162。(PubMed)
- 雅各比,J., A. Nath, Z.F. Jessen和G.W. Schwartz,Amacrine细胞的自我调节缝隙连接网络控制视网膜中一氧化氮的释放。神经元。2018;100 (5): 1149 - 1162. - e5。(PubMed)
- 格莱姆斯,W.N, J.张,C.W.格雷顿,B.卡查尔,J.S.戴蒙德,视网膜并行处理器:100多个独立微电路在单个中间神经元内运行。神经元。2010;65(6): 873 - 885。(PubMed)
- Yadav, S.C, S. Tetenborg和K. Dedek,A8无间隙连接由连接蛋白36组成,但其调控方式与全细胞无间隙连接不同。分子神经科学前沿,2019;12:99。(PubMed)
- 达西,D。感知的起源:视网膜神经节细胞多样性和平行视觉通路的创造,在认知神经科学.2004年,麻省理工学院出版社。(PubMed)
- 桑恩斯,J.R.和R.H.马斯兰德,视网膜神经节细胞的类型:神经元分类的现状和意义。神经科学,2015;38:221 - 246。(PubMed)
- 巴登,P.贝伦斯,K.弗兰克,M. Román Rosón, M.贝其,T.欧拉,小鼠视网膜神经节细胞的功能多样性。大自然。2016;529(7586): 345 - 350。(PubMed)
- Vaney、组长。许多不同类型的视网膜神经元在注射生物素或神经生物素后显示示踪偶联。神经科学信件。1991;125(2): 187 - 190。(PubMed)
- Vaney、组长。兔视网膜定向选择神经节细胞的区域组织。神经科学杂志:神经科学学会官方杂志。1994;14 (11 Pt 1): 6301 - 6316。(PubMed)
- Hidaka, S, Y.赤堀和Y.黑泽明,哺乳动物视网膜神经节细胞之间的树突电突触。《神经科学杂志:美国神经科学学会官方杂志》2004;24(46): 10553 - 10567。(PubMed)
- Müller, l.p.d.s., M.T.H. Do, k - w。邱淑华、何淑华及鲍德里奇小鼠视网膜内感光性视网膜神经节细胞与无分泌细胞的示踪偶联。比较神经学杂志,2010;518(23): 4813 - 4824。(PubMed)
- Reifler, a.n., A.P. Chervenak, M.E. Dolikian, B.A. Benenati, B.Y. Li, R.D. Wachter, A.M.林奇、德默茨、梅耶斯、阿布法哈、耶克尔、弗兰纳里、王家耀,所有在移位的无分泌细胞上持续的尖刺,都接受来自黑视素神经节细胞的间隙连接输入。当代生物学:CB。2015;25(21): 2878。(PubMed)
- 舒伯特,T., S.马克西纳,O. Krüger, K.威勒克和R.威勒,Connexin45介导小鼠视网膜双层神经节细胞的缝隙连接偶联。《比较神经病学杂志》2005;490(1): 29-39。(PubMed)
- Kántor, O, G. Szarka, Z. Benkő, Z. Somogyvári, E. Pálfi, G. Baksa, G. Rácz, R. Nitschke, G. Debertin,和B. Völgyi,人视网膜神经节细胞树突状树突树突中连接36间隙连接的战略定位。细胞神经科学前沿,2018;12:409。(PubMed)
- 姚,X, J.卡法罗,A.J.麦克劳林,F.R.波斯特马,D.L.保罗,G.阿瓦拉马尼,和G.D.菲尔德,缝隙连接有助于通过方向选择性视网膜神经节细胞的差异光适应。神经元。2018;100 (1): 216 - 228. - e6。(PubMed)
- 胡,E.H.和S.A.布卢姆菲尔德,间隙连接耦合下的短潜伏期峰同步视网膜α神经节细胞。《神经科学杂志:美国神经科学学会官方杂志》2003;23日(17):6768 - 6777。(PubMed)
- 特伦霍姆,A.J.麦克劳克林,D.J.施瓦布和G.B.阿瓦拉玛尼,运动编码视网膜神经元网络中电和化学突触传输的动态调谐。神经科学学报:美国神经科学学会官方杂志。2013;33(37): 14927 - 14938。(PubMed)
- 库尔,s和g。w。施瓦茨,原发视网膜神经节细胞通过异型RGC间隙连接接收断开输入,在神经科学学会年会.2019年,神经科学学会:芝加哥。225.08 p。。(PubMed)
- Hu e.h., F. Pan, B. Völgyi, S.A. Bloomfield,光增加视网膜神经节细胞的缝隙连接。《生理学杂志》2010;588 (Pt 21): 4145 - 4163。(PubMed)
- S.H DeVries,兔视网膜神经节细胞的相关放电。神经生理学杂志1999;81(2): 908 - 920。(PubMed)
- 巴洛,h·b, r·菲茨休和s·w·库夫勒,猫在黑暗适应过程中视网膜接受区组织的变化。《生理学杂志》1957;137(3): 338 - 354。(PubMed)
- Peichl L.和H. Wässle,猫视网膜α神经节细胞感受野中心的结构相关性。《生理学杂志》1983;341:309 - 324。(PubMed)
- 穆勒,J.F.和R.F.达歇,兔视网膜α神经节细胞在暗适应下失去拮抗环绕反应。视觉神经科学。1997;14(2): 395 - 401。(PubMed)
- strong, P.K.和F. Rieke,阳伞视网膜神经节细胞间相关活性的起源。自然神经科学杂志。2008;11(11): 1343 - 1351。(PubMed)
- 纳尔逊,R。在猫的视网膜中,所有的大汗腺细胞都能加速杆状信号的时间进程。神经生理学杂志。1982;47(5): 928 - 947。(PubMed)
- Völgyi, B., M.R. Deans, D.L. Paul和S.A. Bloomfield,哺乳动物视网膜中多杆通路的收敛和分离。《神经科学杂志:美国神经科学学会官方杂志》2004;24(49): 11182 - 11192。(PubMed)
- 伊万诺娃,E., U. Müller和H. Wässle,小鼠视网膜双极细胞甘氨酸能输入的特性。《欧洲神经科学杂志》2006;23(2): 350 - 364。(PubMed)
- Münch, t.a., R.A. da Silveira, S. Siegert, T.J. Viney, G.B. watramani和B. Roska,通过多功能神经回路处理的视网膜接近敏感度。自然神经科学杂志。2009;12(10): 1308 - 1316。(PubMed)
- 特雷克斯勒,e·b, w·李,还有S.C.梅西,两根杆状通路对双极细胞和双极细胞光响应的同时贡献。神经生理学杂志。2005;93(3): 1476 - 1485。(PubMed)
- Abd-El-Barr, m.m., M.E. Pennesi, S.M. Saszik, A.J. Barrow, j.m., D.E. Bramblett, D.L. Paul, L.J. Frishman和S.M. Wu,暗适应小鼠视网膜中杆状和锥状通路的遗传解剖。神经生理学杂志。2009;102(3): 1945 - 1955。(PubMed)
- 苏西,E.,王莹,S. Nirenberg, J. nathan, M. Meister,哺乳动物视网膜杆状光感受器到神经节细胞的新信号通路。神经元。1998;21(3): 481 - 493。(PubMed)
- 李文伟、强志伟及梅西,兔视网膜中杆状细胞和OFF锥双极细胞之间的直接突触连接。比较神经学杂志。2004;474(1): 1 - 12。(PubMed)
- 墨菲,G.J.和F. Rieke,神经节细胞的电突触输入是平行视网膜回路输出和绝对敏感性差异的基础。神经科学杂志:神经科学学会官方杂志,2011;31日(34):12218 - 12228。(PubMed)
- 乐意地,G.L。,蟾蜍视网膜中杆状细胞的量子敏感度。科学(纽约,纽约州)。1975;187(4179): 838 - 841。(PubMed)
- 兰姆,T.D.和E.J.西蒙,海龟光感受器细胞间耦合与电噪声的关系。《生理学杂志》1976;263(2): 257 - 286。(PubMed)
- Tessier-Lavigne m.d Attwell,早期视觉加工中光感受器耦合和突触非线性对信噪比的影响。伦敦皇家学会学报。B辑,生物科学,1988;234(1275): 171 - 197。(PubMed)
- 阿特维尔,博尔赫斯,吴世民和威尔逊,信号裁剪杆输出突触。大自然。1987;328(6130): 522 - 524。(PubMed)
- 康纳斯,B.W.和M.A.朗,哺乳动物大脑中的电突触。神经科学,2004;27:393 - 418。(PubMed)
- Pereda, a.e., S. Curti, G. Hoge, R. Cachope, C.E. Flores和J.E. Rash,间隙结介导的电传输:调节机制和可塑性。生物化学与生物物理学报,2013;1828(1): 134 - 146。(PubMed)
- 阿内特,d和t。e。Spraker,兔视网膜神经节细胞维持放电的相互关系分析。《生理学杂志》1981;317:29-47。(PubMed)
- D.N Mastronarde,猫视网膜神经节细胞的相关放电。I.自发主动输入X细胞和y细胞。神经生理学杂志1983;49(2): 303 - 324。(PubMed)
- D.N Mastronarde,猫视网膜神经节细胞的相关放电。2X细胞和y细胞对单个量子事件的响应。神经生理学杂志1983;49(2): 325 - 349。(PubMed)
- D.N Mastronarde,猫视网膜中神经节细胞之间的相互作用。神经生理学杂志1983;49(2): 350 - 365。(PubMed)
- Brivanlou, i.h., D.K. Warland和M. Meister,视网膜神经节细胞协同放电的机制。神经元。1998;20(3): 527 - 539。(PubMed)
- M.格雷什纳,J.施伦斯,C.巴克利察,G.D.菲尔德,J. l .戈瑟,L.H.杰普森,A.谢尔,A.M.Litke和E.J. Chichilnisky,灵长类动物视网膜主要神经节细胞类型的相关放电。《生理学杂志》2011;589 (Pt(1): 75 - 86。(PubMed)
- Völgyi, B., F. Pan, D.L. Paul, J.T. Wang, ad . Huberman, S.A. Bloomfield,缝隙连接对于产生相邻视网膜神经节细胞的相关尖峰活动至关重要。《公共科学图书馆•综合》。2013;8 (7): e69426。(PubMed)
- S. Trenholm, A.J. McLaughlin, D.J. Schwab, M.H. Turner, R.G. Smith, F. Rieke和G.B. Awatramani,电和化学突触输入的非线性树突整合驱动精细尺度的相关性。自然神经科学杂志。2014;17(12): 1759 - 1766。(PubMed)
- D.N Mastronarde,视网膜神经节细胞的相关放电。《神经科学趋势》,1989;12(2): 75 - 80。(PubMed)
- 施尼策,M.J.和m.m Meister,从眼睛到大脑的信号的多神经放电模式。神经元。2003;37(3): 499 - 511。(PubMed)
- 班尼特,M.V.L.和R.S.祖金,哺乳动物大脑中的电耦合和神经元同步。神经元。2004;41(4): 495 - 511。(PubMed)
- 欧施,T.欧拉和W.R.泰勒,兔视网膜的方向选择树突状动作电位。神经元。2005;47(5): 739 - 750。(PubMed)
- 西维尔,B.和S.R.威廉姆斯,方向选择性是通过视网膜神经节细胞中活跃的树突整合来计算的。自然神经科学杂志。2013;16(12): 1848 - 1856。(PubMed)
- 奥伊斯特,C.W.和H.B.巴洛,兔视网膜定向选择单元:首选方向的分布。科学(纽约,纽约州)。1967;155(3764): 841 - 842。(PubMed)
- 徐智,曾强,石旭和,何s,改变出生后早期小鼠视网膜中ON-OFF方向选择性神经节细胞的耦合模式。神经科学。2013;250:798 - 808。(PubMed)
- 乌斯雷,w。m。j。b。雷帕斯和r。c。里德,配对脉冲相互作用和突触效能的视网膜输入丘脑。大自然。1998;395(6700): 384 - 387。(PubMed)
- 卡兰迪尼,J.C.霍顿和L.C.辛奇,通过突触后总和的丘脑对视网膜尖突的过滤。视觉杂志。2007;7(14): -11 - 20.1。(PubMed)
- 克莱兰,b.g., M.W.杜宾和W.R.莱维克,同时记录外侧膝状神经元的输入和输出。《自然:新生物学》1971;231(23): 191 - 192。(PubMed)
- 陈,c。g。雷格尔,视网膜膝状突触的发育重塑。神经元。2000;28(3): 955 - 966。(PubMed)
- 摩根,j.l., D.R. Berger, A.W. Wetzel和J.W. Lichtman,小鼠视丘脑网络连接的模糊逻辑。细胞。2016;165(1): 192 - 206。(PubMed)
- ronpani, S.B, f.e Müllner, A. Wanner, C. Zhang, C. n . Roth, K. Yonehara和B. Roska,单细胞启动的跨突触追踪揭示外侧膝状核视觉整合的不同模式。神经元。2017;93 (4): 767 - 776. - e6。(PubMed)
- 麦斯特,M, l,拉格纳多,和地方检察官贝勒,视网膜神经节细胞的协调信号。科学(纽约,纽约州)。1995;270(5239): 1207 - 1210。(PubMed)
- 肖丽娟,张明,邢东,邢培军。梁和S. Wu,视网膜亮度适应编码策略的转变:从放电速率到神经相关性。神经生理学报。2013;110(8): 1793 - 1803。(PubMed)
- 阿克特,j.m.,吴舜臣,J.C.李,J.艾布拉姆斯,E.H.胡,I.帕尔曼和S.A.布卢姆菲尔德,光诱导的哺乳动物视网膜上偶联ON方向选择神经节细胞之间的穗同步变化。《神经科学杂志:神经科学学会官方杂志》2006;26日(16):4206 - 4215。(PubMed)
- Neuenschwander, S.和W. Singer,猫视网膜和外侧膝状核中振荡光反应的远程同步。大自然。1996;379(6567): 728 - 732。(PubMed)
- Roy K. S. Kumar和S.A. Bloomfield,视网膜大泌腺和神经节细胞之间的间隙连接耦合构成了整体物体感知的整体连贯活动。美国国家科学院学报,2017;114 (48): E10484-E10493。(PubMed)
- Manookin m.b. S.S. Patterson和C.M. Linehan,灵长类视网膜梧桐神经节细胞运动敏感性介导的神经机制。神经元。2018;97 (6): 1327 - 1340. - e4。(PubMed)
- Murphy-Baum, B.L.和G.B. Awatramani,旧神经元学习新技巧:重新定义灵长类视网膜的运动处理。神经元。2018;97(6): 1205 - 1207。(PubMed)
- 贝瑞,m.j., I.H. Brivanlou, T.A. Jordan和M. Meister,视网膜对移动刺激的预期。大自然。1999;398(6725): 334 - 338。(PubMed)
- 约翰斯顿,J.和L.拉格纳多,视网膜连接组的一般特征决定了运动预期的计算。eLife。2015;4.(PubMed)
- 特伦霍尔姆、D.J.施瓦布、V.巴拉苏布拉曼尼亚和G.B.阿瓦拉玛尼,电耦合神经网络的滞后归一化。自然神经科学杂志。2013;16(2): 154 - 156。(PubMed)
- Ball, W.和E. Tronick,婴儿对即将发生的碰撞的反应:视觉和现实。科学(纽约,纽约州)。1971;171(3973): 818 - 820。(PubMed)
- 石根,H., M.甘木,S.本田,和M.立花,同步视网膜振荡编码青蛙逃跑行为的基本信息。自然神经科学杂志。2005;8(8): 1087 - 1095。(PubMed)
- 金,s.m., c·戴克曼,p·雷德格雷夫,p·迪恩,在实验室环境中,利用分散注意力的任务来获得成年人对若隐若现的视觉刺激的防御性头部运动。感知。1992;21(2): 245 - 259。(PubMed)
- 希夫,W, j。a。卡维内斯,j。j。吉布森,恒河猴对“若隐若现”的光学刺激的持续恐惧反应。科学(纽约,纽约州)。1962;136(3520): 982 - 983。(PubMed)
- 特米泽,我,j。c。多诺万,h。拜尔和j。l。赛默哈克,斑马鱼幼虫隐现诱发逃逸的视觉通路。当代生物学:CB。2015;25(14): 1823 - 1834。(PubMed)
- 伊尔马兹,迈斯特先生,小鼠对若隐若现的视觉刺激的快速先天防御反应。当代生物学:CB。2013;23日(20):2011 - 2015。(PubMed)
- 比斯瓦斯,C.哈西列,A.马塔鲁加,G.图曼,P.沃尔特,F. Müller,rd10视网膜固有神经元振荡的药理学分析。《公共科学图书馆•综合》。2014;9 (6): e99075。(PubMed)
- 博洛斯卡,J., S.特伦霍尔姆和G.B.阿瓦拉玛尼,退化小鼠视网膜中的固有神经振荡器。神经科学杂志:神经科学学会官方杂志,2011;31日(13):5000 - 5012。(PubMed)
- 马戈利斯,d。j。g。纽柯克,t。欧拉,p。b。德特维勒,视网膜神经节细胞变性引起的突触输入改变后的功能稳定性。神经科学杂志:神经科学学会官方杂志。2008;28日(25):6526 - 6536。(PubMed)
- J.和G. Zeck,杆状变性小鼠视网膜的网络振荡。神经科学杂志:神经科学学会官方杂志,2011;31日(6):2280 - 2291。(PubMed)
- Stasheff,顺丰速递,视网膜退化(rd1)小鼠神经节细胞出现持续自发多动和OFF反应的暂时保留。神经生理学杂志。2008;99(3): 1408 - 1421。(PubMed)
- Stasheff, s.f., M. Shankar, M.P. Andrews,rd1和rd10小鼠视网膜神经节细胞自发和光诱发活性的变化在发育时间过程中得到区分。神经生理学杂志。2011;105(6): 3002 - 3009。(PubMed)
- Yee, C.W, A.H. Toychiev和B.T. Sagdullaev,网络缺陷加剧视网膜变性小鼠模型的损伤。系统神经科学前沿,2012;6:8。(PubMed)
- Lepore,例如,伴有视力丧失的自发视觉现象:104例视网膜和神经传入通路病变患者。神经学。1990;40 (3 Pt 1): 444 - 447。(PubMed)
- Murtha T.和S.F. Stasheff,视网膜和视神经疾病中的视觉功能障碍。神经诊所。2003;21(2): 445 - 481。(PubMed)
- dr . Weiland和ms . Humayun,视网膜假体。IEEE生物医学工程汇刊。2014;61(5): 1412 - 1424。(PubMed)
- 巴斯坎普,S.皮卡德,J.A.萨赫勒和B.罗斯卡,色素性视网膜炎的光遗传学治疗。基因治疗。2012;19(2): 169 - 175。(PubMed)
- Yee, c.w., A.H. Toychiev, E. Ivanova和B.T. Sagdullaev,视网膜变性小鼠模型视网膜神经节细胞的异常突触输入随形态变化。比较神经病学杂志。2014;522(18): 4085 - 4099。(PubMed)
- 塞肯亚克,C., L.H.杰普森,P.霍托威,A.谢尔,W.达布罗斯基,A.M.Litke和E.J. Chichilnisky,视网膜变性过程中大鼠神经节细胞生理特性的变化。神经生理学杂志。2011;105(5): 2560 - 2571。(PubMed)
- 特伦霍姆,S., J.博罗斯卡,J.张,A.霍加斯,K.约翰逊,S.巴恩斯,T.J.刘易斯,G.B.阿瓦拉玛尼,电耦合AII amacrine-ON锥双极细胞网络中产生的本征振荡活动由电压门控Na+通道驱动。生理学杂志,2012;590(10): 2501 - 2517。(PubMed)
- 伊万诺娃,E., C.W. Yee, R. Baldoni和B.T. Sagdullaev,视网膜变性中的异常活动损害中央视觉处理,依赖于包含cx36的缝隙连接。实验眼研究。2016;150:81 - 89。(PubMed)
- Telias, M., B. Denlinger, Z. Helft, C. Thornton, B. Beckwith-Cohen和R.H. Kramer,视黄酸诱导多动,阻断其受体揭示光反应并增强视网膜变性的视力。神经元。2019;102(3): 574 - 586。e5。(PubMed)
- Choi, H, L. Zhang, M.S. Cembrowski, C.F. Sabottke, A.L. Markowitz, D.A. Butts, W.L. Kath, J.H. Singer和H. Riecke,在rd1小鼠视网膜中,所有大泡细胞的内在爆发是视网膜振荡的基础。神经生理学杂志。2014;112(6): 1491 - 1504。(PubMed)
- 伊万诺娃,E, C.W. Yee和B.T. Sagdullaev,RD视网膜AII无分泌细胞Cx36缝隙连接磷酸化增加。细胞神经科学进展。2015;9:390。(PubMed)
- 库克,J.E.和D.L.贝克尔,视网膜发育中的间隙连接蛋白:“连接”的新角色。生理学(马里兰州贝塞斯达)。2009;24:219 - 230。(PubMed)
- Akopian, A, T. Atlasz, F. Pan, S. Wong, Y. Zhang, B. Völgyi, D.L. Paul, S.A. Bloomfield,缝隙连接介导的视网膜神经元死亡是连接蛋白和损伤特异性的:神经保护的潜在靶点。神经科学杂志:神经科学学会官方杂志。2014;34(32): 10582 - 10591。(PubMed)
- Kranz, F. Paquet-Durand, R. Weiler, U. Janssen-Bienhold和K. Dedek,色素性视网膜炎间隙连接介导的旁观者效应的测试:继发性视锥死亡不因视锥上连接蛋白36的缺失而改变。《公共科学图书馆•综合》。2013;8 (2): e57163。(PubMed)
- V. Paschon, G.S.V. Higa, R.R. Resende, L.R.G. Britto和A.H. Kihara,在机械创伤引起的急性退行性变中,阻断连接蛋白介导的通信促进神经保护。《公共科学图书馆•综合》。2012;7 (9): e45449。(PubMed)
- 斯特里丁格,K., E.彼得拉什-帕韦兹,G.佐德尔,M.纳皮雷,C.迈耶,t.t.埃塞尔,和R.德尔米泽尔,连接蛋白36的丢失增加视网膜细胞对继发性细胞丢失的脆弱性。欧洲神经科学杂志,2005;22(3): 605 - 616。(PubMed)
- 阿罗约,地方检察官,洛杉矶·柯克比和m·b·费勒,视网膜波调节视网膜内光敏神经节细胞回路。The Journal of Neuroscience: The Official Journal of Neuroscience. 2016;36(26): 6892 - 6905。(PubMed)
- 特伦霍姆,s·g·b·阿瓦拉玛尼,第九章-电耦合视网膜网络的动态特性,在网络功能与可塑性荆洁,编者。2017,学术出版社。p . 183 - 208。(PubMed)
- Baldridge W.H。,硬骨水平细胞的三期适应。脑科学进展。2001;131:437 - 449。(PubMed)
- Veruki M.L.和E. Hartveit,电突触介导哺乳动物视网膜杆状通路中的信号传递。神经科学杂志:神经科学学会官方杂志。2002;22日(24):10558 - 10566。(PubMed)
- s.a. Bloomfield, D. Xin和T. Osborne,兔视网膜所有无分泌细胞间耦合的光诱导调节。视觉神经科学。1997;14(3): 565 - 576。(PubMed)
- 舒伯特,J.德根,K.威勒克,S.G.霍尔木兹迪,H.莫耶和R.维勒,Connexin36介导小鼠视网膜α神经节细胞的间隙连接偶联。《比较神经病学杂志》2005;485(3): 191 - 201。(PubMed)
- 拉诺尔,F.和R.A.西尔弗,用间隙连接读取树突活性。自然神经科学杂志。2014;17(12): 1625 - 1627。(PubMed)
- 特伦霍姆,s·g·b·阿瓦拉玛尼,退化视网膜自发活动的起源。细胞神经科学进展。2015;9:277。(PubMed)