马修Schmolesky
人类的视觉系统可以检beplay体育公司测和区分各种各样的刺激,这些刺激可能是彩色的或无色的,可能是动态的或非动态的,可能是有模式的或无模式的,可能是二维的或三维的。值得注意的是,在某种意义上,视觉刺激对视网膜产生的神经最终产物总是相同的。在经历了复杂的光转导、水平细胞和无分泌细胞提供的横向相互作用以及神经节细胞树突整合信号之后,仅剩下沿着神经节细胞轴突传播的不断变化的动作电位流来通知我们的视觉感知。这些看似相同的信号必须在大脑皮层下和皮层中以某种方式进行处理,以创造我们所经历的全方位视觉感知。如何实现这一点是目前占据了成千上万研究人员职业生涯的一个谜题,解决方案的基本框架在过去几十年才开始展开。
为了了解皮层和皮层下的处理过程,我们可以就离开视网膜的信号提出两个基本问题:
- 携带信号的离散解剖通路是什么?
- 这些信号实际上携带了什么信息?
这些问题的答案并不存在。也就是说,到目前为止,我们得到的答案是不完整的,而且在许多情况下是相互矛盾的。我首先指出这一点,是因为读者应该意识到视觉研究是一个新兴的领域,每周都有新的文章发表,带来更多关于大脑如何处理其重大任务的细节。更复杂的是,最常被研究用来理解视觉处理的动物(猴子、猫、雪貂、大鼠、老鼠、仓鼠、斑马鱼、金鱼等)的颅骨中不方便携带同一个大脑的小、中、大、超大拷贝。虽然进化决定了这些物种共有的许多视觉系统设计,但也存在着实质性的差异。beplay体育公司为了这篇综述的目的,我将几乎完全集中在猕猴的视觉系统上,因为它是迄今为止我们对人类视觉最了解和最好的模型之一。beplay体育公司我将从视觉系统研究的历史角度开始,并试图回答上面提出的两个与初级视觉皮层(V1)有关的基本问beplay体育公司题。
历史的角度来看。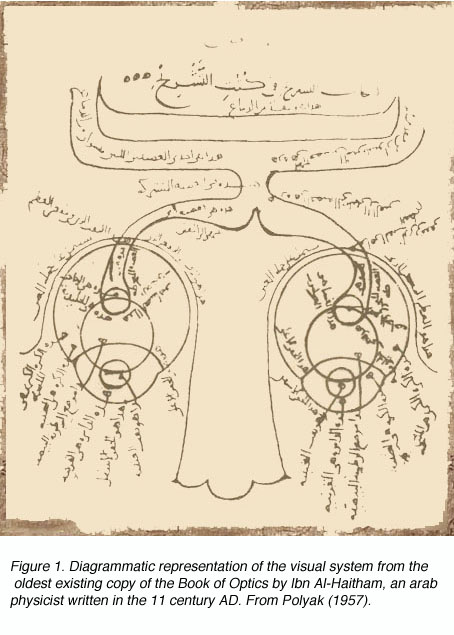
图1。图解表示的视觉系统从现存的最古老的拷贝beplay体育公司《光学是阿拉伯物理学家伊本·阿尔-海瑟姆在公元11世纪写的。从波里亚克(1957)
视觉和眼睛之间的关系,从人类存在的最早时期就必须得到正确的理解。然而,要超越这一最基本的认识,就需要仔细解剖动物和/或人类的眼睛、光纤和大脑结构。一种观点认为,前古典文明(如埃及人、苏美尔人、亚述人、巴比伦人、印度人、中国人、印度人等)的宗教和社会教义不允许这样的剖析。地中海周围更原始的牧民和农民没有这样的禁忌,他们被认为在祭祀仪式中研究过基本的解剖学,尽管不是系统的。这些牧民和农民为他们发现的解剖结构进化出了一种术语。
Polyak(1957)坚持认为这个术语在公元前600-400年被希腊人迅速采用,当时解剖学组织的研究被严格追求。在此期间,希波克拉底(公元前460-380年)、阿尔克迈翁(公元前520年)和安纳萨哥拉斯(公元前500年)等医生对动物和人类的身体和大脑进行了解剖,并对大脑功能(如运动、感觉和思考)持有一定的准确性意见。
希腊医学科学知识的高度是在公元前323年至公元前212年的希腊化或亚历山大时期达到的。最多产的医学家是佩加蒙(今土耳其)的“内科王子”盖伦(公元129-201年)。正如Polyak(1957)所指出的,盖伦对理解正常和病理条件下的人体解剖和生理学的贡献是巨大的,并为阿拉伯时期的医学奠定了基础,随后为中世纪后期和近代欧洲的复兴奠定了基础。图1显示了阿拉伯人在从眼睛到前脑的视觉系统通路上对盖伦思想的适应。
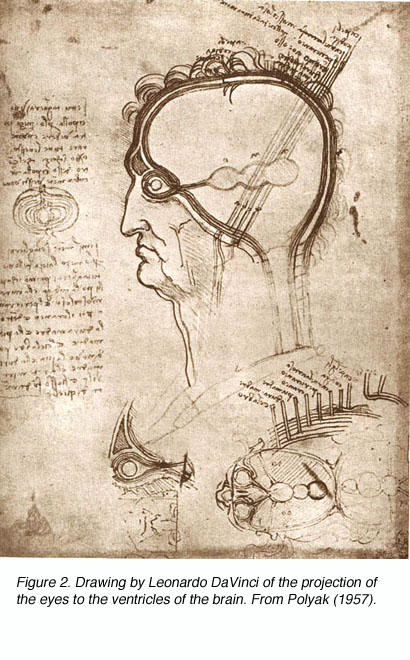
虽然Galen准确地描述了视觉系统的大体解剖结构,但他没有识别出视交叉处的纤维交叉,也没有追beplay体育公司踪到丘脑的背侧外侧膝关节状核(LGNd)(图1)。相反,他认为这些纤维连接到侧脑室(他将其命名为“丘脑”)。这一观点与当时关于神经的普遍观点是一致的,即神经是承载各种“精神”的空心通道,如视觉精神。一千多年后,人文主义科学家如列奥纳多·达·芬奇(他描绘了同样的光纤到侧脑室的通路)(图2)和德·卡特(他提倡神经携带动物精神的观点)的信仰附和了盖伦的学说,并证明了在如此长的时间内取得的进展是多么的微小。
在希腊化时期之后,基督教时代最初几个世纪所支持的伦理学否定了医生和科学家解剖人类尸体的权利。这种狭窄在黑暗时代被扩大到包括解剖动物,从这一点到17世纪,治疗师被迫几乎完全依赖他们从古典时期收集到的少量信息。
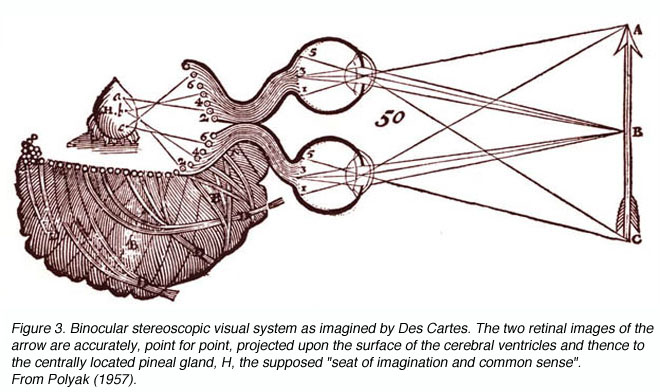
图3。德·卡特设想的双目立体视觉系统。beplay体育公司箭头的两个视网膜图像精确地,点对点地投射在脑室表面,并从那里投射到位于中央的松果体H,这被认为是想象力和常识的中心。从波里亚克(1957)
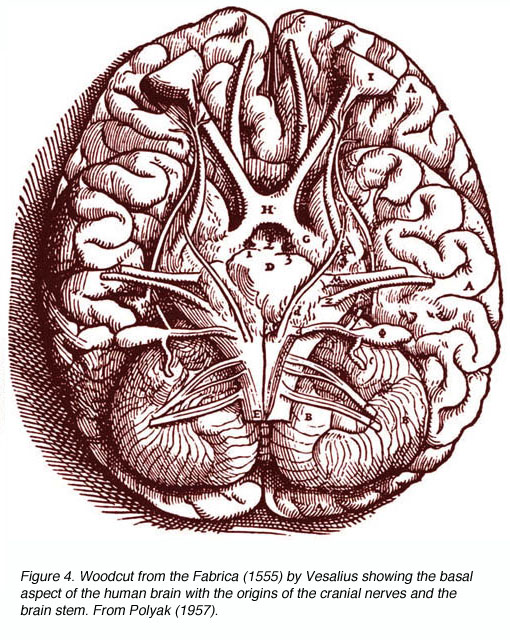
图4。木刻的以(1555)维萨利乌斯所作,展示了人类大脑的基础部分,其中包括颅神经和脑干的起源。从波里亚克(1957)
在公元1600年到1860年之间,人们对中央视觉系统的解剖和功能有了最基本的了解。beplay体育公司推导了光纤在交叉处的部分交叉。这些纤维投射到LGNd和光学辐射到皮质被描述。意外和实验诱导的皮质病变分别与患者和实验动物的症状相关,导致了局部皮质功能的概念。仔细进行皮质解剖,Gennari线(密集的有髓纤维束穿过V1层4B;以它的发现者命名)在初级视觉皮层中被描述。
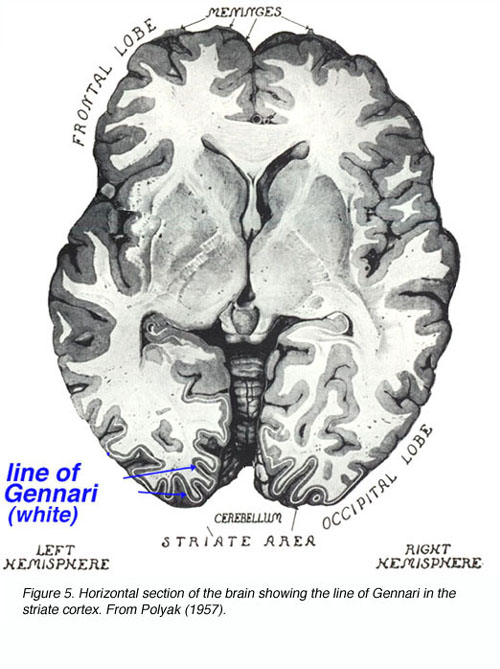
图5。大脑横切面显示条纹皮层的Gennari线。从波里亚克(1957)
这种条纹产生了术语“纹状皮层”来描述V1。在玻片上检查未染色的皮层部分,向baillager显示皮层的6层。中心结构视网膜位组织的概念得到了广泛的接受。
在19世纪末和20世纪初,组织学准备方面的几项进展导致了视觉皮层细胞研究的繁荣。在切薄片之前,人们发现了更好的化学治疗方法来硬化大脑;开发了总神经元和纤维染色染色方案(如威格特和尼赛尔的方案);建立了标记单个细胞的高尔基技术及其广泛的轴突和树突树突。
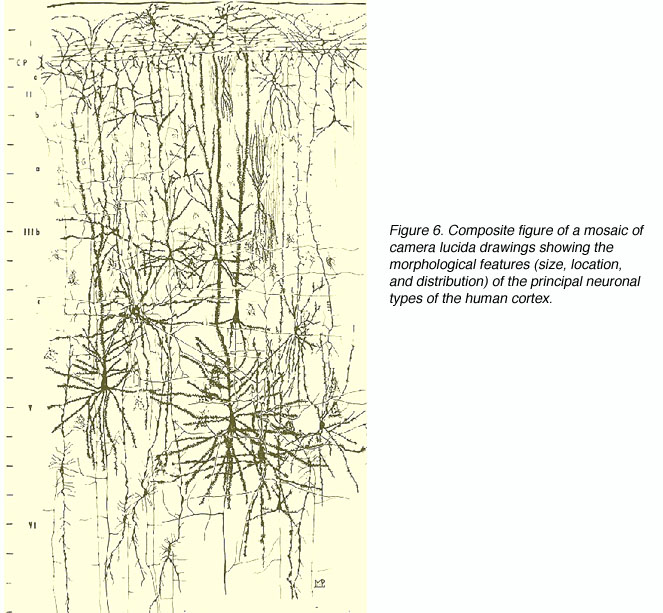
图6。摄像头lucida绘制的马赛克复合图,显示了人类皮层主要神经元类型的形态特征(大小、位置和分布)。(来自:rapid-Golgi准备。规模:100µm)
研究人员第一次可以看到神经元形态的复杂性和层流差异的细节。在这方面,没有一个研究者比圣地亚哥Ramón y Cajal(1852-1934)贡献更大。Ramón y Cajal使用高尔基法和他自己改进的银染色法详细描述了整个皮层(视觉和其他)的细胞形态。这些技术提供的令人难以置信的细节使Ramón y Cajal提出了某些神经通路和回路,并对皮层成分的功能意义进行了理论化。电子显微镜的出现使研究人员能够描述突触连接的细节。“示踪剂”染料被远端纤维吸收并运输到细胞体(逆行示踪剂)或被细胞/树突吸收并沿着纤维运输到远端末端(顺行示踪剂)的开发极大地增强了视觉系统中前馈和反馈途径的搜索(细胞核和/或纤维束病变后的特异性跨突触变性曾被使用,但收效甚微)。beplay体育公司
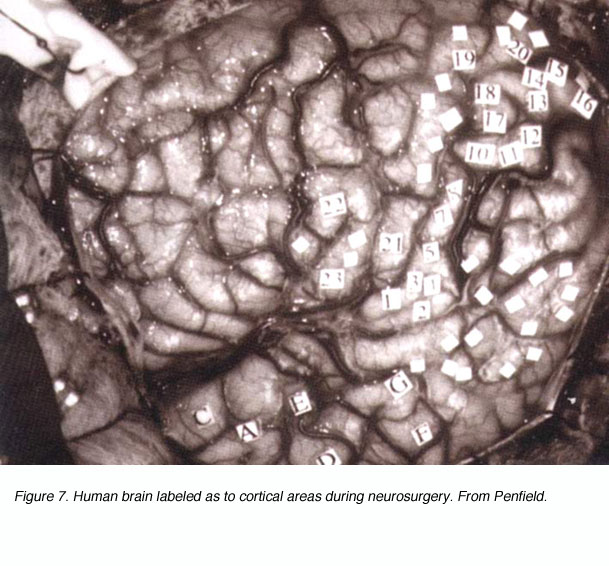
以上所述的所有进展在本质上都是解剖学的。那视觉通路的功能呢?在19世纪末和20世纪初,研究人员是如何处理这个问题的?几个世纪以来描述的临床病例已经指出了分散的皮层区域的功能专门化。皮层内的实验病变证实了这一观点。对大脑皮层的电刺激(如Hitzig和Fritsch对狗和猴子运动皮层的电刺激)为功能对等学说提供了致命一击,并建立了取而代之的大脑功能定位理论。彭菲尔德等外科医生在手术过程中对意识清醒的患者的大脑进行机械刺激,并记录所唤起的感觉(图7)。然而,当研究人员从电刺激转向电记录时,获得视觉系统功能图的关键步骤就到来了。beplay体育公司
使用视觉诱发电位(VEPs)的头皮记录来绘制视觉反应皮层区域,并检查兔、猫和猴的视网膜位组织和双眼视功能(Talbot和Marshall, 1941;汤普森等人,1950年;丹尼尔和怀特里奇,1959年)。同时,微电极被用于研究视网膜/视道中的单细胞/纤维(或小群)(例如,Hartline, 1938;Kuffler, 1953)和体感皮层(Mountcastle, 1957;鲍威尔和蒙特卡斯尔,1959年)。Hubel和Wiesel首次将单单元记录技术应用于猫(1959年,1962年)和猴子(1968年,1974年,1977年)的视觉皮层,结果令人震惊。随着这一系列令人难以置信的实验(David Hubel和Torston Wiesel后来因此获得了1982年的诺贝尔奖),视觉皮层研究的现代时代开始了。
考虑到这一历史观点,我们应该将注意力转向目前对初级视觉皮层及其在视觉通路中的位置的理解。
基本的解剖学。
和所有被研究的哺乳动物一样,猕猴的初级视觉皮层位于枕叶皮层的后极(图8)。
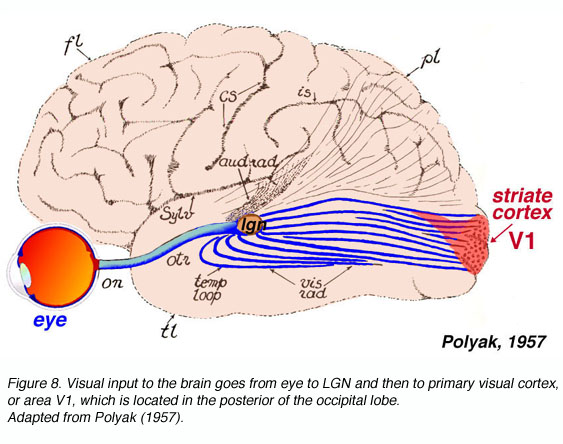
图8。大脑的视觉输入从眼睛到LGN,然后到位于枕叶后部的初级视觉皮层或V1区。改编自《波利亚克》(1957)
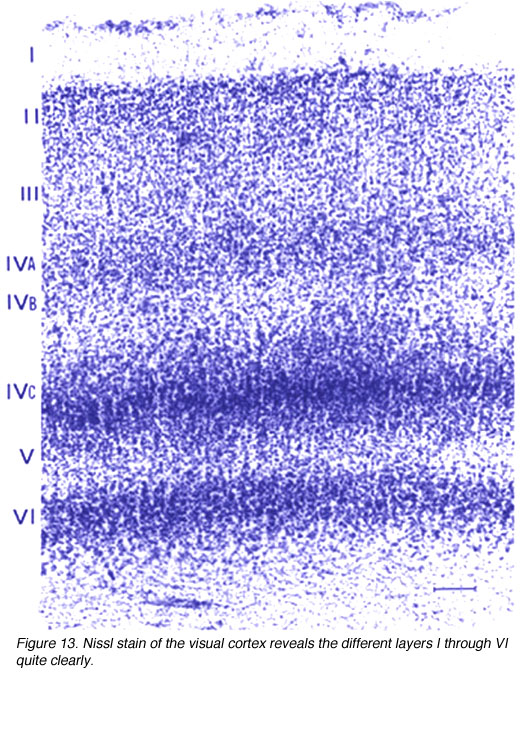
V1向外侧几乎延伸到月骨沟,后外侧几乎延伸到枕下沟;V1/V2边界在任何沟槽之前相遇。大部分V1存在于钙质沟内,V1在这些褶皱中的界限可以通过V1第4层的明显出现来标识(图9)。
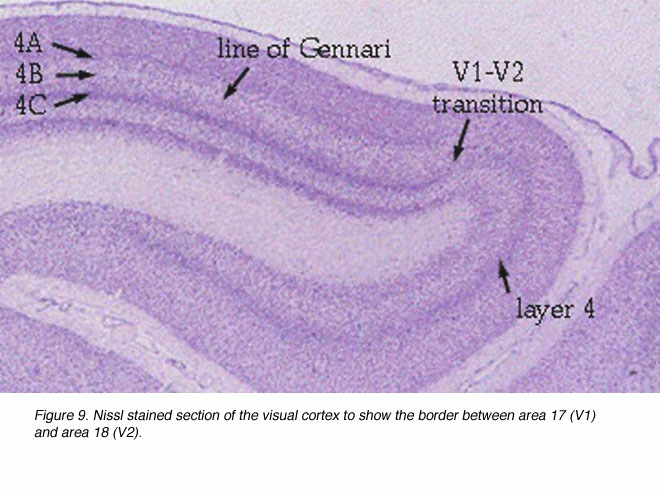
图9。尼氏染色切片显示17区(V1)和18区(V2)之间的边界
灵长类动物初级视觉皮层的解剖结构已经得到了详尽的研究。灵长类动物V1的三个基本组织原则是:(1)兴奋和抑制神经元的层层状和(2)柱状排列(与其他新皮层区域共享的排列)和(3)细胞色素氧化酶(CO)标记显示的解剖/功能隔室的规则间距。
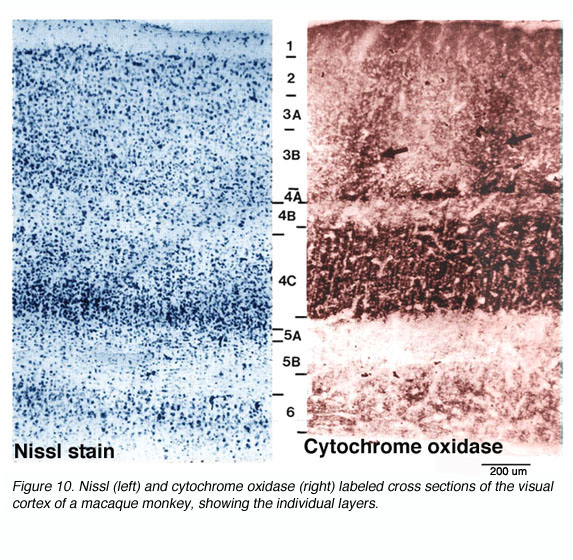
图10。Nissl(左)和细胞色素氧化酶(右)标记的猕猴视觉皮层横截面,显示了单个层
在处理这些解剖特征的细节之前,可以陈述几个简单的事实和经验法则:
* Brodmann介绍的6个皮层层(第1层最靠背,第6层最靠腹)随着神经元的其他方面及其连接被揭示,被一再细分
*第1层几乎动脉瘤样,主要由树突和轴突连接组成。在一毫米三的V1组织中,可以发现大约4700个神经元,2900个小胶质细胞,3400个少突胶质细胞和49000个星形胶质细胞(O’kusky和Colonnier, 1984)。存在的神经元中超过80%是gaba能神经元(Fitzpatrick et al., 1987)。
* 2-6层中大约20%的神经元是抑制性中间神经元(GABAergic),它们对V1电路的功能起主要作用,但不会将轴突投射到该区域之外。
* 2/3层(“超颗粒”层)包含许多兴奋性投射神经元,将轴突发送到纹外皮层区域(如V2、V3、V4、MT等)。
*第4层(颗粒层)分为4个水平子层:4A, 4B, 4C一个, 4 cb.层4摄氏度一个和4 cb是LGNd神经支配的主要接受者。LGNd的大细胞层(M)和细小细胞层(P)投射到4C一个和4摄氏度b,分别。因此,M和P流在这个阶段仍然是分离的(但请参阅下面的限定条件)。
* 5/6层(“粒内”层)包含许多兴奋性投射神经元,支配LGNd向这个中继区域提供反馈。
*对V1和V2中CO含量的标记显示了V1中称为“斑点”(或泡、斑点或斑块)和“斑点间”的CO丰富和贫乏区域,以及V2中的厚、薄和淡条纹。这些分类的相关性仍然是一个有争议的问题。
* CO斑点不均匀地从第1层表面延伸到第6层底部。相反,它们集中在3B、4B和4C层(图10)。
*到CO斑点和间斑点的投影相对不同如下:
一)Pb(灵长类P细胞或小神经节细胞)投射到LGNd细小细胞层(参见视网膜小通路章节),投射到V1层4C,然后投射到V1层2/3的间团,最后这些层投射到V2膝状肌的浅条纹
b)蓝黄神经节细胞(参见S锥通路和色盲章节)投射到LGNd的插层和S/K (K为“koniocellular”)层,投射到V1层的斑点上,2/3投射到V2层的细条纹上
P c)一个(M细胞或阳伞神经节细胞)投射到LGNd的大细胞层,再从那里投射到V1的4C层。这一层投射到V1的2/3层和V1的4B层。4B层依次投射到中颞区(MT)和V2的粗条纹。
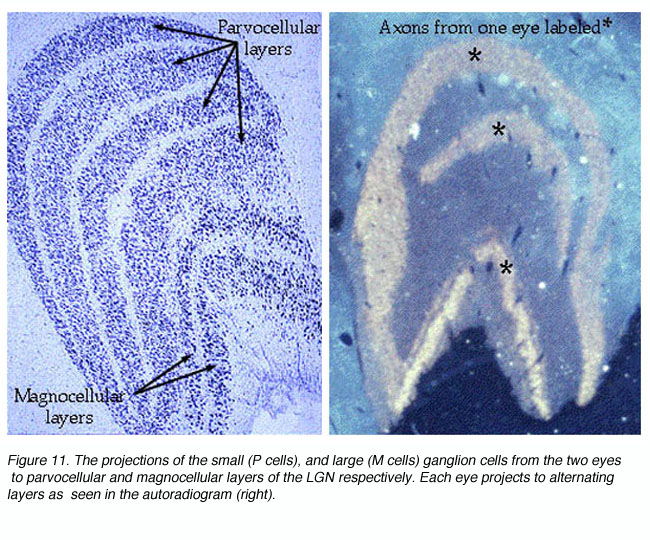
图11。两眼小(P细胞)和大(M细胞)神经节细胞分别投射到LGN的细小细胞层和巨细胞层
*第1层和第2层接收来自纹外皮层的反馈输入。
*幼年成年猕猴V1的神经胶质细胞占所有细胞的20-35%(不包括周细胞)。神经胶质细胞可分为少突胶质细胞(约占细胞的7-10%)、星形胶质细胞(约占细胞的11-20%)和小胶质细胞(约占细胞的1.5-3%)(O’kusky and Colonnier, 1982;彼得斯等人,1991)。
神经元的选民。
灵长类动物V1中有三种基本类型的神经元(图12):
-
- 多刺锥体细胞(兴奋性)
-
- 刺状星状细胞(兴奋性)
- 光滑的或稀疏的棘状中间神经元(几乎都是GABAergic)。
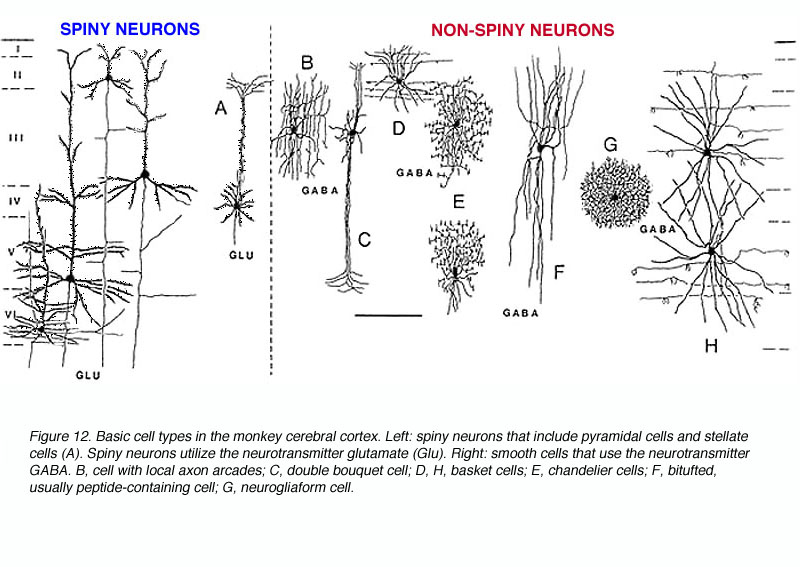
如你所料,锥体细胞的细胞体呈金字塔状,顶端有树突(即向白质延伸),树突上覆盖着刺。带刺的星状细胞通常较小,它们的细胞体更像一个星形。它们也有刺状树突(图12A)。中间神经元的细胞体更圆,树突上几乎没有刺。多年来,解剖学家们或多或少地对一些独特的子类型作出了描述性的命名,下面简要地描述一下。
- 吊灯细胞抑制;轴突表现出类似烛台的垂直分支(图12E)。
- Neurogliaform细胞抑制;类似神经胶质细胞但绝对是神经元的;轴突和树突分支,但保留在体细胞内,形成致密的球状纤维区(图12G)。
- 双束细胞抑制;轴突侧支和支突树突垂直延伸成紧密的束(图12C)。calbindin抗体专门标记这些细胞,使用这种策略,研究人员发现这些细胞在第3层有规律地间隔,从而形成另一个柱状单位。每10000个花束细胞中大约有7-15个双花束细胞米米2(DeFelipe等,1990)。
- 篮子里的细胞(Somogyi et al, 1983;DeFelipe等人,1986)-抑制;较大的细胞体向外侧延伸有髓神经轴突分支很远。轴突分枝以类似篮子的方式终止于金字塔状的细胞体和树突上。树突垂直延伸(图12D,H)。
- Cajal-Retzius细胞——抑制;Cajal和Retzius分别在新生动物和人类的第一层细胞中独立发现了这种细胞类型。它们可能存在,也可能不存在于成人体内。
- Meynert细胞——兴奋性;由Meynert首次描述的大锥体细胞;“外部”Meynert细胞体在4B中发现,相对于锥体细胞锥有规律地分布(Peters和Sethares, 1991;见下文)。这些细胞投射到MT区。“内部”Meynert细胞体在5/6层边界有规律地间隔,位于CO间斑的下方。这些细胞也存在于第六层。这些细胞典型地具有不对称的、横向的树突状树状化,具有高度的方向选择性,也可投射到MT区。
上面描述的细胞类型仅代表了少数比较独特的变种。詹妮弗·隆德及其同事(隆德,1987;Lund等人,1988;隆德和吉冈,1991年;Lund和Wu, 1997)使用高尔基浸渍染色法对猕猴V1的推定中间神经元进行了严格的研究。基于一系列形态学标准(体体的层流位置、轴突树突、树枝状树突树突、分支的模式和程度),他们单独描述了40多种中间神经元。
其他低密度中间神经元可能仍未识别。根据化学成分和/或生理反应特性,可以进一步划分已描述的类(图12)。或者,应该注意到,许多解剖学上截然不同的类型在生理学上彼此之间可能无法区分。
皮质层。
一般来说,新皮层层有许多编号方案,特别是V1层。我采用了布罗德曼的基本方案,他根据仔细的组织学定义了皮层层。我还采纳了Lund和同事的工作建议(见下文)。
第1层由2-5B层锥体细胞的顶端树突(Lund and Wu, 1997)和LGNd K层收集的输入之间形成的密集突触网络组成(Fitzpatrick et al., 1983;Lachica和Casagrande, 1992),枕区,来自纹外区、非特异性丘脑核和其他皮层下区域的反馈通路。因此,虽然第1层神经元很少,但它是一个网络层,对更深层锥体细胞的放电特性有直接一致的影响。
粒上层(2,3a和3B)含有大量锥体细胞的体块和树突。第2层和第3A层收到的丘脑输入很少(K层投射到V1 blob主要集中在3B)。它们不直接接收来自4C层的输入,而3B层则接收大量的4C输入。第2层和第3A层确实包含许多在所有其他皮层层中发现的神经元轴突和树突。
第4A层在尼氏染色中显示为小颗粒状细胞的深色带。这一点和锥体细胞的缺乏将这一层与3B层区分开来。
4B层是细胞贫乏层,它包含低密度排列的锥体细胞和大的外层Meynert细胞。这一层从底层4C接收强大的输入一个Meynert细胞可以直接投射到MT区和上丘(SC;《炸薯条与Distel》,1983年)。4B层的细胞也投射到V2的厚CO条纹上,而V2又投射到MT上。因此,该层在M通路/背侧流的信息传递中起着重要作用。
4C层与4B层的区别部分在于该层中有大量的星形细胞堆积。波利亚克(1957)把4个辛分为一个而且b单纯基于组织学的分类;上面的4C神经元更加分散。膝状神经支配模式的差异(即LGNd到4C的M层)一个P层到4C层b;Hubel和Wiesel, 1972;Blasdel and Lund, 1983)、4C细胞的投射模式(Lund, 1973)和生理特性(Blasdel and Fitzpatrick, 1984)都压倒性地支持了这种划分。更多的证据表明,4C的上部和下部一个也可能是不同的(Blasdel and Lund, 1983;隆德,1987;Yabuta和Callaway, 1998)。
4摄氏度b
多刺的星形神经元在4A和3B层有密集的神经支配,在4C、5和可能6层有稀疏的神经支配(Lund, 1973)。没有来自4C的直接预测b已发现3A、4B或5B (Lund, 1987)。相比之下,如前所述,4C一个严重依赖于4B。
局部5A层神经元不投射到4B,只弱地支配4Cb.它们支配着所有其他层。5A神经元似乎主要投射到3B和1(Lund, 1987)。5B中的锥体神经元将循环轴突发送到3A层(Lund and Boothe, 1975)。5/6边界和第6层内的Meynert细胞投射到MT和SC (Fries等人,1985年)。
第6层:已知第6层的神经元将循环轴突发送到4Cb(隆德和布斯,1975)。其他神经元(甚至可能是相同的神经元)将轴突投射回LGNd (Lund et al., 1975)。第6层还从LGNd接收直接输入(Hendrickson等,1978),从而形成一个神经环路。
V1皮质列。
术语“皮层柱”指的是从皮层表面到白质垂直排列的细胞可能包括功能或解剖学单位。因此,皮层柱可以根据解剖学特征(如锥体细胞顶端树突束的定型模式)、功能特征(如皮层细胞柱均对同一刺激方向作出反应)或两者来定义。
许多类型的列被提出,包括眼优势,方向,空间频率和颜色列。这些柱状排列的细节将在后面讨论V1生理学的章节中描述。然而,我将在这里简要描述眼优势柱,以提供一个在初级视觉皮层柱状排列的例子。
眼优势列。
来自两只眼睛的视觉信号在LGNd(图11)和V1区域的geniculoreceiver层保持分离(图14)。我们可以通过测量4C层单元的电生理反应来观察这种分离。当记录电极在4C层内移动时,有一个突然的移动,即哪只眼睛驱动单元。在4C层,从一只眼睛到另一只眼睛的转换发生在不到50微米的距离内。来自这些波段的信号汇聚到皮层浅层的单个神经元上,从而形成由一只眼睛或另一只眼睛交替支配的柱状结构。
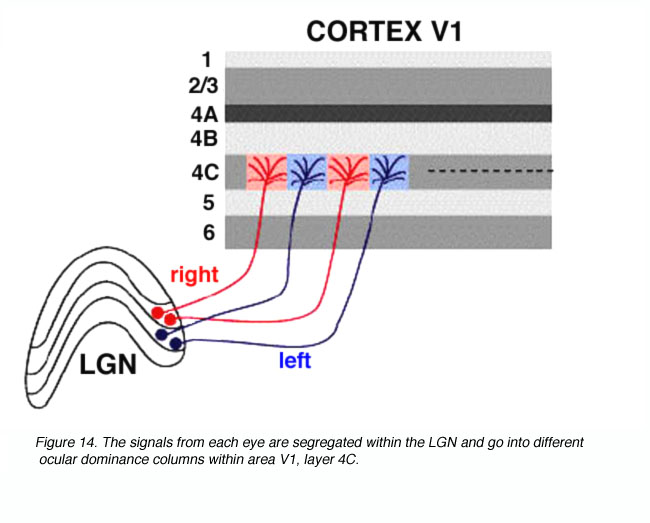
图14。来自每只眼睛的信号被分离到V1区域,4C层的不同的眼优势柱中
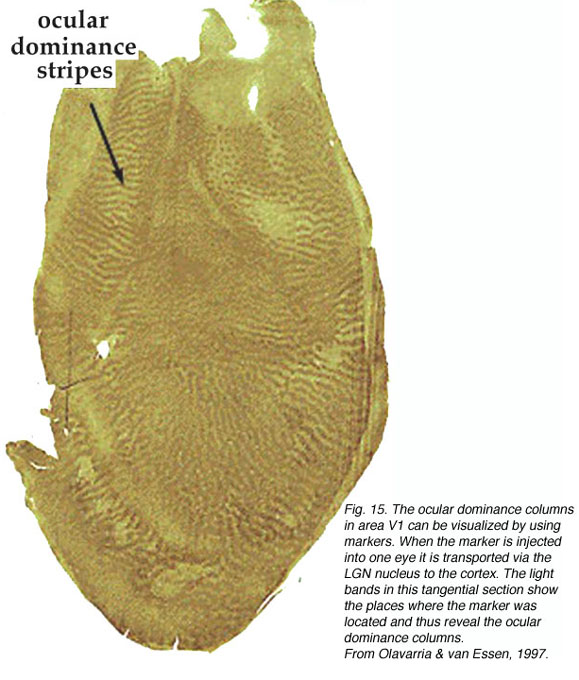
细胞色素氧化酶标签。
细胞色素氧化酶(CO)是一种完整的跨膜蛋白,存在于线粒体内膜。因为它的作用是催化三磷酸腺苷(ATP)的产生,而ATP本身是一种能量分子,在大量的细胞过程中使用,所以细胞或细胞核中CO的数量可以作为细胞活性的指标。1978年,Margaret Wong-Riley发现,除了将CO作为一种活动指标外,初级视觉皮层的CO标记还揭示了一系列显著的CO富区和缺区(即斑点和斑点间区)(图16)。
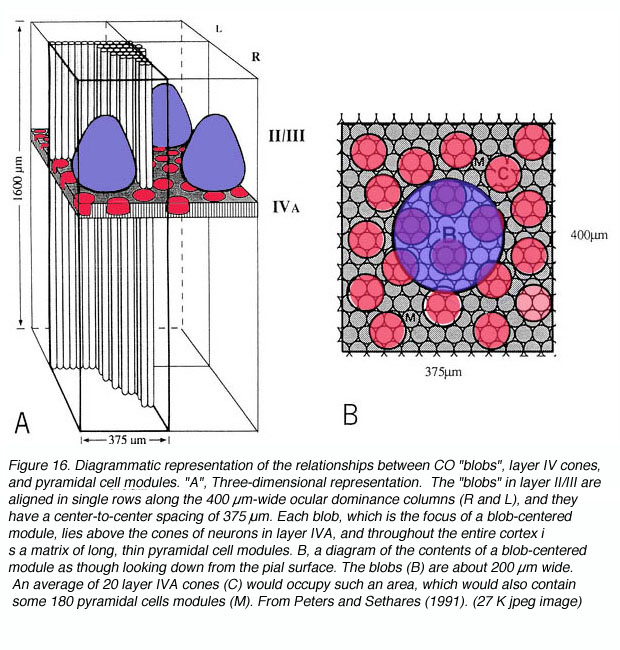
据推测,这一区域反映了两组以前未被区分的神经元在能量消耗方面的稳定差异。假设这是正确的,那么blobs和interblobs之间还可能存在什么其他差异呢?首先,应该注意的是,致密的CO染色并不存在于整个皮质深度。相反,CO斑点在3B、4A和4C层最明显。尽管如此,如果这些密集区域实际上在某种程度上与缺co的相邻区域不同,那么富裕和贫穷区域的正上方和正下方的细胞也可能表现出与它们上方和下方细胞相关的连通性和功能特征的差异。
例如,3B层的斑点已被证明与猕猴特定的膝关节神经支配相对应(Livingstone和Hubel, 1982;亨德利和吉冈,1994)。Boyd和Casagrande(1999)最近发现,在灌木幼猴和猫头鹰猴中,投射到MT的V1细胞在3B层CO斑点下方比在inter斑点下方数量更多。猕猴的类似数据也显示,通过V1的横向区域,MT投射细胞以周期性步骤聚集在一起(Shipp和Zeki, 1989),但没有发现这些聚集与CO之间的相关性。
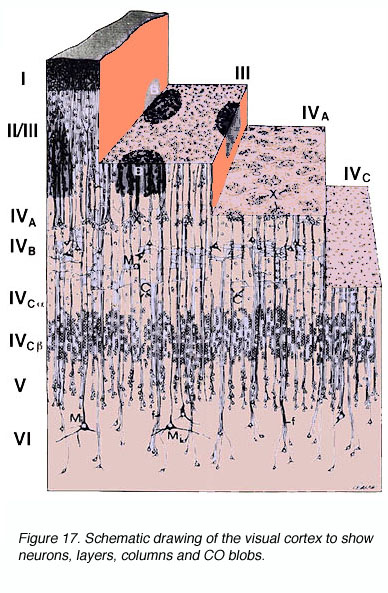
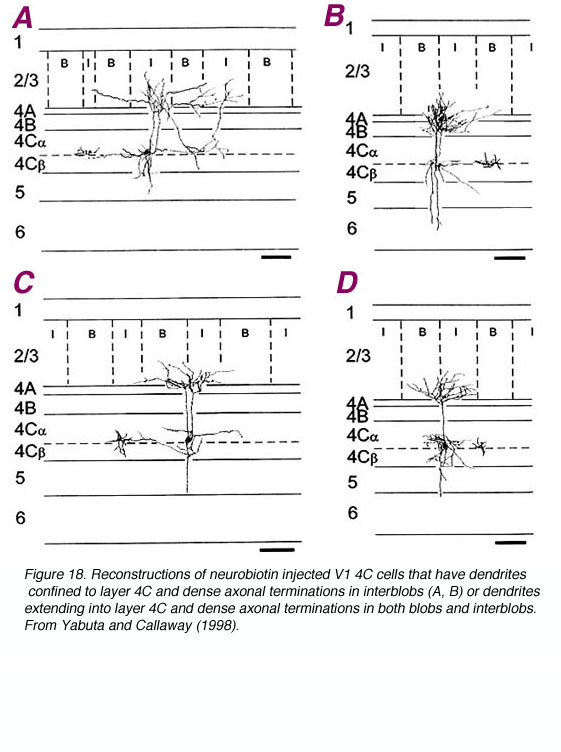
图18。注入神经生物素的V1 4C细胞,其树突局限于4C层,在斑点间(A, B)中有密集的轴突末端,或树突延伸到4C层,斑点和斑点间均有密集的轴突末端。选自Yabuta和Callaway (1998)
前馈和反馈路径。
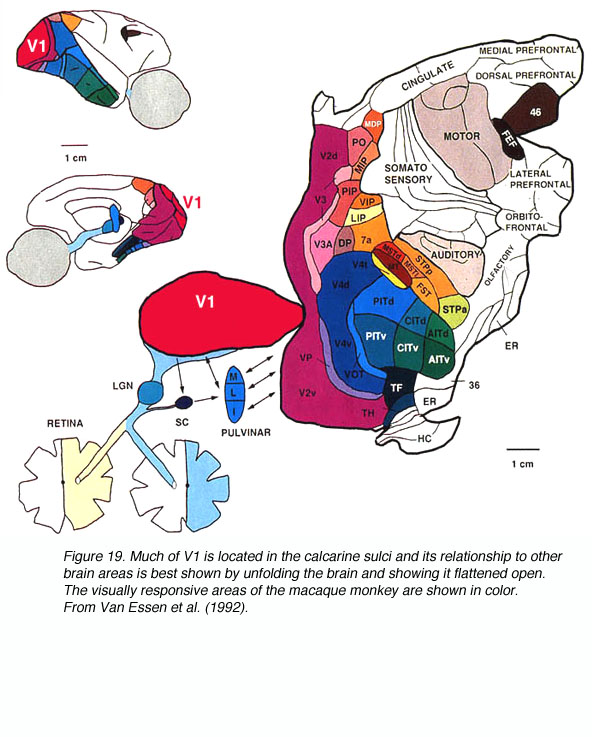
来自两个视网膜的信号通过LGN传递到V1区。在猕猴中,在V1中处理信号后,它们通过多条通路传递到30多个视觉响应的纹外皮层区域(图19)。
{kind=link}
{kind=link}
{kind=link}
图19所示。V1的大部分位于钙质沟中,它与大脑其他区域的关系最好的展示是展开大脑并将其摊开。猕猴的视觉反应区域以彩色显示。来自Van Essen等人(1992)
解剖学层次模型将视觉皮层区域放入一个多层处理模型中,该模型基于在每个区域中发现的前馈、横向和反馈路径的模式。一般来说,前馈通路被认为是从一个区域的粒上层投射到目标区域的第4层。相反,反馈投影是从一个区域的粒内层产生的,并在目标区域的第4层外终止(见Felleman和Van Essen, 1991;Hilgetag等人,1996;Barone等人,2000年)。
在前面的章节中,我已经描述了V1的解剖边界,存在于该区域的细胞,这些细胞在层、柱和CO隔室中的排列,我已经暗示了进入和离开V1的视觉通路。的确,视觉信息从视网膜传递到LGNd到V1到更高的皮层区域,而更高的皮层区域投射回V1和V1到LGNd(灵长类动物的视网膜没有传出投射)。然而,尽管这种视觉处理的观点是准确的,但它没有捕捉到V1中发现的大量上下游连接。
对V1的直接前馈投射来自于Meynert的枕部、LGNd、闭锁、中心旁核、中缝系统、蓝斑核和基底核(Ogren and Hendrickson, 1976;雷扎克和贝内文托,1979年;格雷厄姆,1982;布拉斯德尔和隆德,1983年;多提,1983;Perkel等人,1986;拉奇卡和卡萨格兰德,1992年;亨德利和吉冈,1994;亚当斯等人,2000年)。
从V1直接前馈投影延伸到V2、V3、V5或MT、MST和FEF (Lund等人,1975;Maunsell和Van Essen, 1983;Ungerleider和Desimone 1986a,b;利文斯通和胡贝尔,1987年;希普和泽基,1989年;Boussaoud等,1990;Fitzpatrick等人,1994)。
对V1的直接反馈投射来自于V2、V3、V4、V5或MT、MST、FEF、LIP和颞下皮层(Ungerleider and Desimone, 1986a,b;Perkel等人,1986;希普和泽基,1989年;罗克兰等人,1994;Barone等人,2000年;铃木等人,2000年。
V1的直接反馈投射延伸到SC、LGNd、枕区和桥脑(Fries和Distel, 1983;隆德等,1975年;格雷厄姆,1982;薯条,1990;古铁雷斯和Cusick;1997)。
上面列出的连接只是将信号带进带出初级视觉皮层的直接和间接投射的一个子集,甚至还没有开始描述V1内部广泛的横向连接。
视觉信号携带什么信息?
我们现在知道了V1的层、柱和CO室中神经元、树突和轴突的排列。我们知道许多进出V1的通路涉及视网膜、下皮层和纹状外皮层。我们可以想象光线照射到眼睛上,从这个原点开始,一连串动作电位从一个神经元到另一个神经元,从一个区域到另一个区域,向前,横向,再向后。抽象地说,我们知道正是这些信号决定了我们对外部世界的看法。但这数以百万计的图案尖刺是如何分解成一个实际的知觉的呢?一个部分的答案是,考虑到每张图像本身都可以被分解成组成部分(如线条、颜色、纹理、阴影和运动等),视觉皮层神经元专门用于检测这些组成部分的子集。
接受域的属性。
视觉皮层中的每个细胞都有一个感受野,这是一个相对于中央凹的空间上的离散区域,在这里视觉刺激的呈现或移除都会引起细胞激活。根据定义,这个感受野之外的刺激既不会增加也不会减少该单个细胞的持续活动。感受野(RF)的位置和大小可以被认为是射频最基本的特性之一
V1中的视网膜位图。
视网膜内神经节细胞的空间位置是由LGN层内神经元的空间组织所保留的。LGN后方含有感受野位于中央凹附近的神经元。感受野的位置从后向前发展,逐渐成为视网膜的外围(见Erwin et al., 1999)。这种空间布局被称为视网膜位组织,因为LGN中感受野的拓扑组织与视网膜的组织平行。
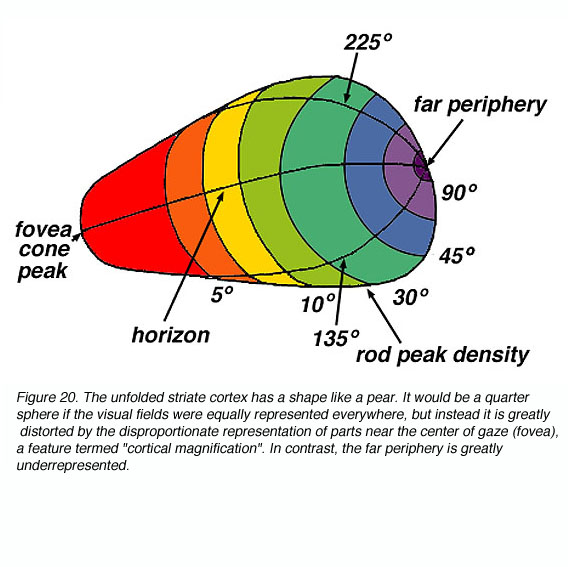
V1区域的信号也按视网膜位置排列。从猴子的电生理学中,我们可以用一个电极测量感受野的位置,电极切向穿过4C层,穿过眼优势柱。沿着这条路径的神经元的感受野中心系统地对应着从中央凹到外周的位置。这种趋势在局部被眼优势边界处的小而突然的跳跃打断。因此,纹状体皮层保留了LGN发育的对侧视野的视网膜位图。
定向和方向选择性。
V1是第一个在猕猴身上观察到强烈的定向和方向选择性的位点(Hubel and Wiesel, 1968)。虽然绝大多数V1细胞表现出一定程度的方向选择性,但只有大约25-35%的V1细胞具有强方向选择性(Schiller等,1976;DeValois等人,1982年)。测试定向和方向选择性的经典方法是测量单个单元响应漂移定向亮度条和/或漂移亮度点时的峰值速率(参见图21)。
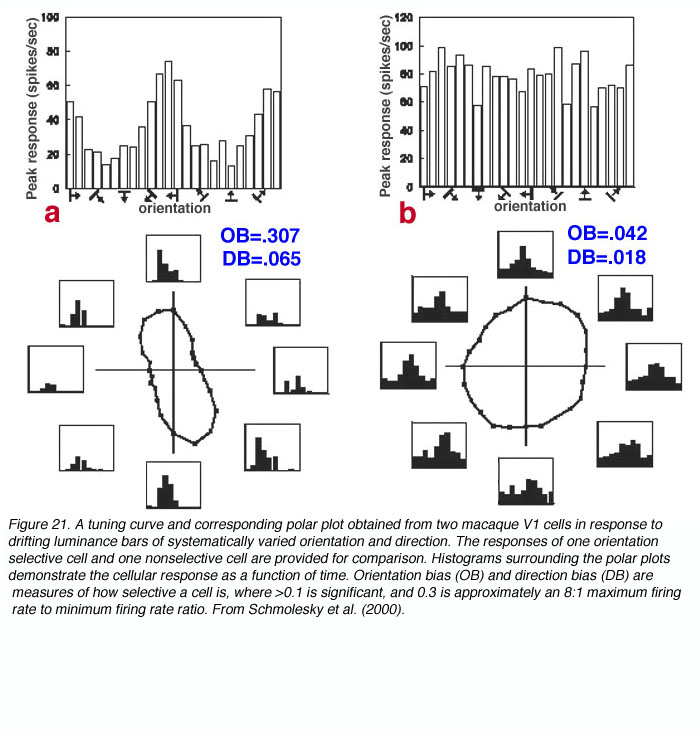
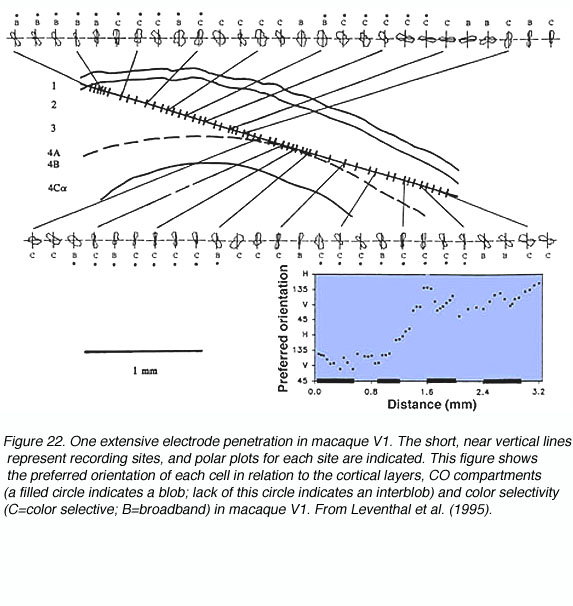
通过查看图20和图21,可以很容易地理解方向列的概念。当电极以与皮层层相对平行的角度降低到V1时(见图22),遇到的细胞的定向选择系统地变化,相邻的细胞区域共享近似的定向偏好。
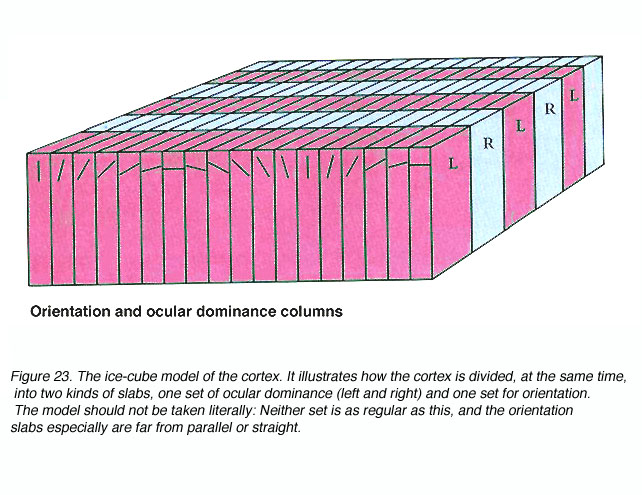
这样的记录使得Hubel和Wiesel提出了如下所示的功能性组织模型(图23)。
最近,最初由Hubel和Wiesel描述的V1的方向柱也被重新定义为更复杂的几何图形,如部分柱(“平板”)和风车,这是由于越来越多的证据(例如Bonhoeffer和Grinvald, 1991)所决定的。
双眼视和双眼视差。
Hubel和Wiesel(1962)首次证明了灵长类动物V1中存在对两只眼睛的视觉刺激有优先反应的细胞,而不是只对一只眼睛有反应的细胞。他们使用7分制对细胞进行分类,其中1 =仅受对侧眼刺激的细胞,2 =对侧刺激强烈偏倚的细胞,3 =对侧刺激弱偏倚的细胞,4 =对双眼刺激反应最大的细胞,5 =对同侧刺激反应弱偏倚的细胞,6 =同侧刺激强烈偏倚的细胞,7 =仅受同侧眼刺激的细胞。单眼细胞(1、2、6、7)与双眼细胞(3、4、5)的比例随细胞所记录层的不同而不同。视网膜缩孔受体层4C细胞一个, 4 cb均为单眼。此后的解剖学研究表明,这些细胞从LGNd接收单眼输入,而LGNd不从交替层汇聚(Hubel和Wiesel, 1972;隆德和布斯,1975)。
双目细胞的用途是什么?当观察者注视一个视觉物体时,该物体的图像定位在两个视网膜的相应区域上。然而,与此同时,注视点前后的物体产生的图像位于两个视网膜的不对应区域上。图像不对应的程度(如通过视网膜偏心的差异评分来衡量)被定义为双眼视差。利用双眼视差来确定一个物体与自己的距离,以及它与注视面的关系的能力被称为立体视。
Hubel和Wiesel首先描述了V1中对双眼视差敏感的神经元的存在。也就是说,当刺激具有正视差和弱甚至抑制(低于基线)到0视差刺激(在固定面)或负视差刺激时,一些神经元可能表现出最大的放电速率。这些细胞被描述为视差敏感,负视差调谐。如果调整相当尖锐,细胞可能被描述为负视差选择性,即它只反应,而不是简单地优先,负视差。我们现在知道了视差敏感细胞的百分比以及正视差、负视差和0视差调谐细胞的比例。显然,在4C层中没有视差调谐细胞一个, 4 cb因为双眼视能力是视差敏感度的先决条件。正如Livingstone和Tsao(1998)所指出的,可以有把握地假设,负的和正的视差选择细胞参与了立体视。在0视差调谐细胞中,所有细胞都可能参与立体视觉,或者,可能只有那些调谐非常窄的细胞(即选择性细胞)有助于这种感知能力。DeAngelis, Newsome和Cumming的工作(DeAngelis et al., 1998;DeAngelis和Newsome, 1999)巧妙地解决了这个问题,他们试图通过微刺激MT区差异调节细胞的子区域来改变训练过的猴子的立体视觉。
视觉通路中的响应时间。
视觉诱发的单细胞反应的另一个特征是时间,目前正受到关注。视觉区域被激活的顺序和任何给定区域内的激活时间范围都是重要的问题。如上所述,灵长目动物视觉的主流模型提出了基于解剖连接性的视觉区域的分层阶段(Felleman和Van Essen, 1991;范·埃森等人,1992年;DeYoe等,1994;Barone等人,2000年)。毫无疑问,一个单一的刺激会以分级的方式启动从视网膜到LGNd到V1的神经元放电特征流。不幸的是,灵长类动物视觉流中皮层连接和功能分化的复杂性使得进一步的预测具有很强的推测性(见Hilgetag et al., 1996)。
例如,我们可以假设,从V1到中颞区(MT)的重髓鞘化路径会导致该区域的快速激活。然而,是所有的MT细胞都以这种方式被激活,还是其他细胞会通过更间接的V1到V2到MT途径等待激活?许多研究报告了单一视觉区域的反应起始延迟。然而,使用这些数据来构建视觉系统激活时间的图表,揭示了一系列的复杂性(Nowak和Bullier, 1998)。beplay体育公司
首先,绝大多数报告延迟的研究只针对一两个视觉区域。因此,10个视觉区域的激活分析可能涉及5-10个独立研究之间的交叉比较。在这样的比较中,我们可以得出这样的结论:平均而言,V1细胞比额视野区(FEF)的细胞活跃得早得多(Maunsell and Gibson, 1992;汤普森等人,1996)。在另一个这样的比较中,我们将被迫得出相反的结论(诺瓦克等人,1995;参考诺瓦克和布利耶,1998年)。
造成这种表面差异的原因,以及许多类似的原因,无疑是使用了不同的实验和数据分析协议。使用的视觉刺激(闪烁vs移动;点、条、格栅或其他;低亮度与高亮度或对比度,等等),使用的动物制剂(麻醉与否),以及标记潜伏期所采取的数据分析策略(定性或定量,等等),在现有的报告中都是不同的。
因此,在最近的一项研究中,我们试图通过使用相同的实验和分析技术从许多灵长类动物的视觉区域获得起病潜伏期来消除这些混淆(Schmolesky et al., 1998)。结果表明,灵长类动物视觉系统的两个主要功能流(M/背侧流参与运动感知和跟踪,P/腹侧流参与物体识别和颜色编码)对时间过程的响应非常不同(见图24)。beplay体育公司
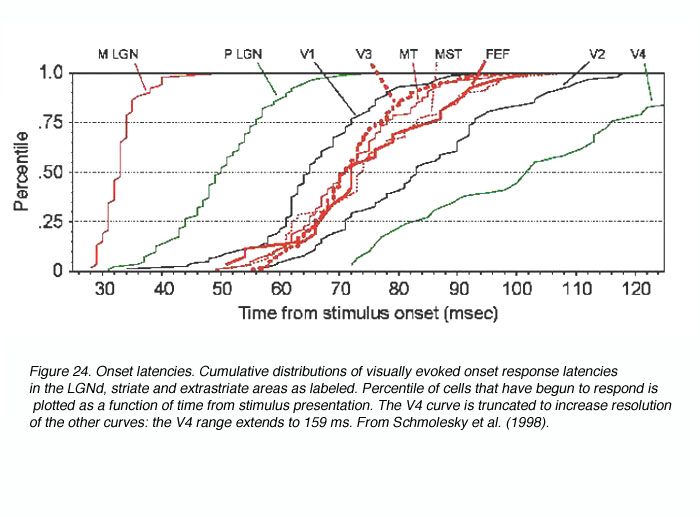
一般来说,M流区域响应迅速而同时,而P流对应区域响应缓慢而依次。特别与这篇综述相关的是,LGNd M细胞变得活跃的速度比P细胞的对应细胞快15-20毫秒(Schmolesky et al., 1998;Maunsell et al., 1999),这种时间分离维持在geniculo受体层4C一个, 4 cb(Maunsell和Gibson, 1992;诺瓦克等人,1995;Schmolesky et al., 1998)。因此,反应时间上的功能分离存在于皮层下和V1的M和P通路中,并似乎被传递到纹外皮层的背侧和腹侧流。
虚幻的轮廓感知。
现代的视觉知觉观是一种动态过程,它超越了提供给视网膜的视觉信息的简单复制。80多年来,格式塔心理学家一直认为,知觉行为创造了格式塔,一种不是被观察对象属性的图形或形式,但代表了大脑对感觉的组织(Kandel, 1991, p441)。这种动态性被认为对完成简单的日常视觉任务(如识别部分遮挡的物体)至关重要。因此,研究大脑如何能够填补缺失的部分是一个重要的课题;这通常是通过使用³虚幻等高线²来实现的(图25)。
错觉轮廓(ICs)是由它们在图像本身没有任何亮度差异的情况下的主观外观所定义的。IC的一个经典例子是虚幻的正方形,其中四个扇形圆盘以适当的方式定向产生正方形的感觉。在这种情况下,很明显,没有真正的亮度差异存在,形成一个完整的正方形,但一个正方形被感知到(图25)。

行为测试表明猫和猴子能够感知这种轮廓(Bravo et al., 1988;De Weerd等人,1990,1996)。最近的生理学研究检查了条纹状和纹外皮层中的单个细胞是否对传统亮度轮廓和幻觉定义的亮度轮廓有相似的反应(例如,Grosof等人,1993;彼得汉斯和冯·德·海特,1989年;冯·德·海特等人,1984;Von der Heydt和Peterhans, 1989)。在每一项研究中,都报道了IC反应细胞。然而,这些研究对IC反应的起源提供了相互矛盾的证据,支持或反对在V1中存在IC反应细胞。不同研究中使用的替代集成电路进一步混淆了这个问题。
我们试图从更广泛的轮廓感知问题(Leventhal et al., 1998)来探讨错觉轮廓感知。这里的问题是,当定义刺激边界的视觉线索(如颜色、亮度、纹理、运动、幻觉等)变化时,大脑如何编码刺激边界(如正方形)。正如格式塔心理学家会注意到的那样,每一种情况下的刺激都是非常不同的,但基本的知觉,正方形,是相同的。我们问了两个基本问题。首先,猫和猴子体内是否存在V1或V2细胞能够以类似的方式对边界做出反应,而不管边界是由什么线索定义的?替代方法包括视觉处理后期的线索不变性的起源,或者通过不同的细胞亚群(每个细胞负责一个线索)对线索信息进行单独的传导。第二,如果这样的细胞存在,它们在V1和V2中是否同样普遍?我们的结果表明,两个区域的细胞亚群可以以线索不变的方式响应刺激边界,如定向棒,尽管这种特性在V1中很少,而在V2中普遍存在。
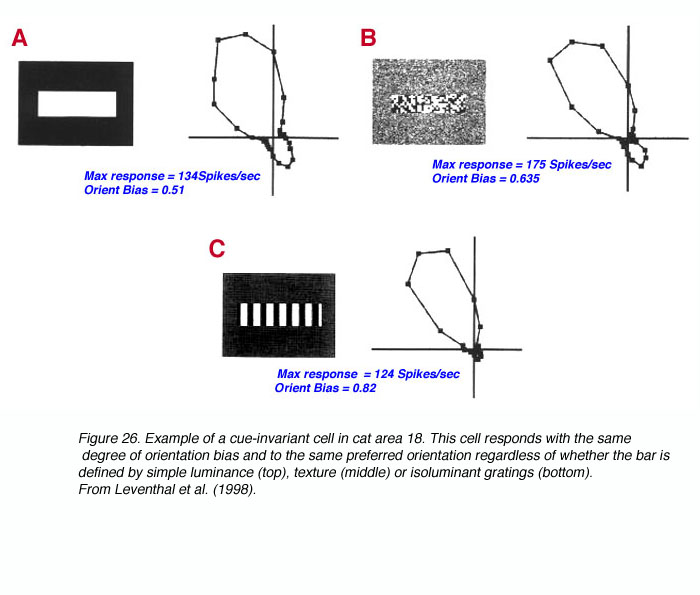
这项工作得出的一个结论是,处于皮层处理第一阶段的细胞已经能够以复杂的方式对单个刺激边界做出反应,即使是在定义物体的线索发生变化或部分闭塞的情况下,也能够检测到物体(Leventhal et al., 1998)。
理解愿景:逆向工程的一个问题。
我们试图理解视觉通路就像接近一台机器,除了它的基本功能外,我们对它一无所知。打个比方,我们都知道如何操作汽车(或多或少),并认识到它的基本功能,把我们从一个地方送到另一个地方。现在让我们假设我们除了这些一无所知,出于某种原因,决定我们应该了解汽车内部工作的每一个细节。我们可以先试着弄清楚汽车的哪些部分专门负责这个核心功能,哪些不是。我们拆除保险杠,喇叭,空调和挡风玻璃,发现汽车仍然运行出色。我们将引擎的小部件一块一块地取出,直到汽车无法启动,类似于对大脑损伤的研究。慢慢地,我们开始理解汽车(大脑)的哪些部分与运动(视觉)有关。在视觉研究中,我们还处在游戏的这个阶段,仍然想知道每个部分实际上是做什么的,什么时候做的,以及所有单独的部分是如何协同工作的。包括我在内的许多研究人员都希望,通过使用化学、电生理、基因和行为方法对视觉通路的结构和功能进行仔细研究,最终能够真正理解大脑是如何为我们提供这种最关键的感官能力——视觉的。
参考文献
亚当斯MM, Hof PR,加塔斯R,韦伯斯特MJ, Ungerleider LG。猕猴枕部视觉皮质投射与化学结构。J Comp Neurol.2000; 419:377 - 393。[PubMed]
Barone P, Batardiere A, Knoblauch K, Kennedy H.投影到视觉区V1和V4的纹外区神经元的层流分布与等级等级相关,表明距离规则的操作。J > 2000; 20:3263 - 3281。[PubMed]
Blasdel GG, Lund JS。猕猴纹状皮层传入轴突的终止。J > 1983; 3:1389 - 1413。[PubMed]
猕猴纹状皮层第4层的生理组织。J > 1984; 4:880 - 895。[PubMed]
猫视皮层的等向畴排列呈针轮状。大自然。1991;353:429 - 431。[PubMed]
Boussaoud D, Ungerleider LG, Desimone R.运动分析路径:猕猴颞内侧上视区和颞上视区眼底的皮层连接。中华神经科学杂志1990;296:462-495。[PubMed]
初级视觉皮层(V1)细胞色素氧化酶(CO)斑点与投射到中颞区(MT)的神经元分布之间的关系。中华神经科学杂志1999;409:573-591。[PubMed]
布拉沃M,布莱克R,莫里森s猫看到主观轮廓。视觉研究》1988;28:861 - 865。[PubMed]
老卡哈尔,脊椎的视网膜。在:Thorpe SA, Glickstein M,翻译。视网膜的结构斯普林菲尔德(伊利诺伊州):查尔斯·c·托马斯出版社;1933.
卡尔Weigert(1845 - 1904)。《美国医学协会杂志》上。1964; 189:769 - 770。[PubMed]
狒狒和猴子的钙质皮层的视野表征。杂志。1959;148:33P。
DeAngelis GC, Cumming BG, Newsome WT。皮质面积MT和立体深度知觉。大自然。1998;394:677 - 680。[PubMed]
陈晓明,陈晓明。猕猴脑区差异选择神经元的组织。神经科学杂志1999;19:1398-1415。[PubMed]
德菲利佩J, Hendry SHC, Hashikawa T, Molinari M, Jones EG。双花束细胞轴突免疫细胞化学研究揭示的猴子大脑皮层微柱状结构。神经科学。1990;37:655 - 673。[PubMed]
德菲利佩,徐亨德利,琼斯等。猴感觉运动皮层篮子细胞和大gaba能神经元的相关电镜研究。神经科学。1986; 17:991 - 1009。[PubMed]
王晓燕,王晓燕,王晓燕。猕猴视觉皮层细胞的定向和方向选择性。视觉研究》1982;22:531 - 544。[PubMed]
De Weerd P, Vandenbussche E, De Bruyn B, Orban GA。猫的错觉轮廓定向辨别。behavior Brain Res. 1990; 39:1-17。[PubMed]
De Weerd P, Desimone R, Ungerleider LG。猕猴V4损伤后光栅定向识别的线索依赖缺陷。Vis > 1996; 13:529 - 538。[PubMed]
DeYoe EA, Felleman DJ, Van Essen DC, McClendon E.枕颞视皮层的多重加工流。大自然。1994;371:151 - 154。[PubMed]
Doty RW光碟。猕猴纹状皮层的非膝状传入。中华神经科学杂志1983;218:159-173。[PubMed]
Erwin E, Baker FH, Busen WF, Malpeli JG。恒河猴外侧膝状核层状拓扑和视网膜拓扑之间的关系:来自功能性图谱的结果。中华神经科学杂志1999;407:92-102。[PubMed]
Essen DC, Van Anderson CH, fellleman DJ。灵长类视觉系统中的信息处理:一个综合系统的视角。beplay体育公司科学。1992;255:419 - 423。PubMed]
范埃森特区的DJ伙计。灵长类动物大脑皮层的分布等级处理。Cereb皮层。1991;1:1-47。[PubMed]
Fitzpatrick D, Itoh K, Diamond IT。松鼠猴(松鼠猴)外侧膝状体和纹状皮层的层状组织。J Neurosci.1983; 3:673 - 702。[PubMed]
Fitzpatrick D, Lund JS, Schmechel DE, Towles AC.猕猴纹状皮层gaba能神经元和轴突末梢的分布。J Comp Neurol.1987; 264:73 - 91。[PubMed]
Fitzpatrick D, Usrey WM, Schofield BR, Einstein G.猕猴纹状皮层第6层皮质合状神经元的层下组织。Vis Neurosci.1994; 11:307 - 315。[PubMed]
弗朗茨·尼塞尔(1860-1919),神经病理学家。《美国医学协会杂志》上。1968; 205:460 - 461。
Fries W, Distel H.猴子纹状体皮层(Meynert细胞)的大V1层神经元投射到上丘。生物学报,1983;219:53-59 .[PubMed]
猕猴纹状皮层(Meynert细胞)的大层VI细胞同时投射到上丘和纹前视区V5。Exp Brain res 1985; 58:613-616。[PubMed]
猕猴条纹状和条纹前视皮层的Fries W.桥脑投影:一项顺行研究。Vis > 1990; 4:205 - 216。[PubMed]
Fritsch G, Hitzig E. Ueber die elektrische Erregbarkeit des Grosshirns。大脑的电兴奋性。《皮毛解剖学、生理学与科学》1870;37:300-332。
猕猴纹状皮层与皮层下结构的一些地形联系。Exp Brain res 1982; 47:1-14。[PubMed]
格罗夫DH,沙普利RM,霍肯MJ。猕猴的V1神经元可以发出“虚幻的”轮廓信号。大自然。1993;365:550 - 552。[PubMed]
古铁雷斯C,库西克CG。猕猴的V1区投射到下枕核复合体的多个组织化学定义的细分区。大脑Res.1997; 765:349 - 356。[PubMed]
Hartline港元。脊椎动物眼睛的单一视神经纤维对视网膜光照的反应。医学杂志。1938;121:400-415。
猕猴膝状背外侧核中神经化学上独特的第三通道。科学。1994;264:575 - 577。[PubMed]
亨德里克森AE,威尔逊JR,奥格伦议员。旧半球和新世界灵长类背外侧膝状核与视觉皮层之间通路的神经解剖学组织。中华神经科学杂志1978;182:123-136。[PubMed]
希尔格塔格CC,奥尼尔MA,年轻议员。视觉系统的不确定的组织beplay体育公司科学。1996;271:776 - 777。[PubMed]
Hubel DH, Wiesel TN.猫纹状皮层中单个神经元的接受野。杂志。1959;150:91 - 104。
猫视觉皮层的感受野、双眼相互作用和功能结构。杂志。1962;160:106 - 154。[PubMed]
猴纹状皮层的感受野与功能结构。杂志。1968;195:215 - 243。[PubMed] [免费全文在PMC]
猕猴膝皮层纤维的层状和柱状分布。中华神经科学杂志1972;146:421-450。[PubMed]
猴纹状皮层定向柱的序列规律性和几何结构。中华神经科学杂志1974;158:267-293。[PubMed]
Hubel DH, Wiesel TN, Stryker MP。用2-脱氧葡萄糖放射自显影技术显示猕猴视觉皮层的定向柱。1977; 269:328 - 330。[PubMed]
休博尔DH。初级视觉皮层的探索,1955-78年。大自然。1982;299:515 - 524。[PubMed]
坎德尔ER。知觉:运动、深度和形状的知觉作者:Kandel ER, Schwartz JH, Jessel TM,编辑。神经科学原理。诺沃克(CT): Appleton & Lange;1991.
Kuffler西南。哺乳动物视网膜放电模式与功能组织。J Neurophysiol。1953;16:37 - 68。[PubMed]
灵长类动物视觉皮层细胞色素氧化酶(CO)斑点的直接w样膝状突起:轴突形态。J Comp Neurol.1992; 319:141 - 158。[PubMed]
李文涛AG,汤臣KG,刘东,周勇,奥尔特SJ。对猴子纹状皮层第2、3和4层细胞的方向、方向和颜色伴随的敏感性。J > 1995; 15:1808 - 1818。[PubMed]
李文涛,王永春,周勇。边界知觉的神经相关研究。Vis > 1998; 15:1107 - 1118。[PubMed]
Livingstone MS, Hubel DH。丘脑输入到猴子视觉皮层富含细胞色素氧化酶的区域。美国国家科学与技术委员会1982;79:6098-6101。[PubMed] [免费全文在PMC]
Livingstone MS, Hubel DH。松鼠猴17区4B层与18区厚细胞色素氧化酶条带之间的联系。J Neurosci.1987; 7:3371 - 3377。[PubMed]
猕猴V1的方向选择机制。神经元。1998;20:509 - 526。[PubMed]
隆德JS。猴(猕猴)视觉皮层第17区神经元的组织。中华神经科学杂志1973;147:455-496。[PubMed]
Lund JS, Boothe RG。猕猴视觉皮层第17区层间连接和锥体神经元组织。J Comp Neurol.1975; 159:305 - 334。
Lund JS, Lund RD, Hendrickson AE, Bunt AH, Fuchs AF。山辣根过氧化物酶逆行转运显示猕猴初级视觉皮层(第17区)传出通路的起源。中华神经科学杂志1975;164:287-303。[PubMed]
隆德JS。猕猴纹状皮层局部回路神经元。一、4C、5A层神经元。中华神经科学杂志1987;257:60-92。[PubMed]
Lund JS, Hawken MJ, Parker AJ。猕猴纹状皮层局部回路神经元。25B和6层神经元。中华神经科学杂志1988;276:1-29 .[PubMed]
猕猴纹状皮层局部回路神经元。3纹层4B, 4A和3B的神经元。中华神经科学杂志。1991;311:234-258 .[PubMed]
隆德JS,吴昌强。猕猴纹状皮层局部回路神经元。四、1-3A椎板神经元。中华神经科学杂志1997;384:109-126。[PubMed]
Maunsell JH, Van Essen DC。猕猴中颞视区(MT)的连接及其与皮层层次的关系。J > 1983; 3:2563 - 2586。[PubMed]
猕猴纹状皮层的视觉反应潜伏期。J Neurophysiol。1992;68:1332 - 1344。[PubMed]
Maunsell JH, Ghose GM, Assad JA, McAdams CJ, Boudreau CE, Noerager BD.猕猴大细胞和小细胞LGN神经元的视觉反应潜伏期。Vis > 1999; 16:1-14。[PubMed]
Mountcastle VB。猫体感觉皮层单个神经元的形态和地形特征。J Neurophysiol。1957;20:408 - 434。[PubMed]
猕猴V1和V2区视觉潜伏期的研究。Vis > 1995; 12:371 - 384。[PubMed]
王晓东,王晓东。视觉系统中信息传递的时序。beplay体育公司编辑:Kaas JH, Rockland K, Peters A。大脑皮层。纽约:全会出版社;1998.p . 205 - 241。
毛猴和沙猴的纹状皮层和皮层下区域之间的通路:枕区相互连接的证据。实验神经。1976;53:780 - 800。[PubMed]
O 'Kusky J, Colonnier M.成年猕猴视觉皮层(第17区)神经元、胶质细胞和突触数量的层流分析。J Comp Neurol.1982; 210:278 - 290。[PubMed]
Olavarria JF, Van Essen DC。猕猴视觉区V2细胞色素氧化酶条带的全局格局。Cereb皮层。1997;7:395 - 404。PubMed]
电刺激对人大脑皮层的躯体运动和感觉表征的研究。大脑。1937;60:389 - 443。
猕猴17区传入连通性的地形:一项双标记研究。J Comp Neurol.1986; 253:374 - 402。[PubMed]
猕猴初级视觉皮层锥体神经元的组织。中华神经科学杂志1991;306:1-23。[PubMed]
孙晓燕,张晓燕。猴子视觉皮层轮廓感知机制的研究。2轮廓弥合差距。J > 1989; 9:1749 - 1763。[PubMed]
脊椎动物的视觉系统。beplay体育公司芝加哥:芝加哥大学出版社,1957
鲍威尔TP,蒙特卡斯尔VB。猴子中央后回皮层功能组织的一些方面:在单个单元分析中获得的结果与细胞结构的相关性。布尔·约翰·霍普金斯医院,1959;105:133-162。[PubMed]
Rezak M,贝内文托洛杉矶。猕猴膝状外侧核背侧、枕下及邻近枕侧侧与初级视觉皮层(第17区)的投射组织比较。大脑研究》1979;167:19-40。[PubMed]
猕猴初级视觉皮层定向调节的动力学研究。大自然。1997;387:281 - 284。[PubMed]
Rockland KS, Saleem KS, Tanaka K.来自猕猴V4区和TEO的发散反馈连接。Vis > 1994; 11:579 - 600。[PubMed]
席勒PH,芬利BL,沃尔曼SF。猴纹状皮层单细胞特性的定量研究。一、感受野的时空组织。J Neurophysiol。1976;39:1288 - 1319。[PubMed]
Schmolesky MT, Wang YC, Hanes DP, Thompson KG, Leutgeb S, Schall JD, Leventhal AG。猕猴视觉系统的信号定时。beplay体育公司J Neurophysiol.1998; 79:3272 - 3278。[PubMed]
王毅,蒲敏,李文涛股份有限公司。衰老恒河猴视觉皮层细胞刺激选择性的退化。Nat > 2000; 3:384 - 390。[PubMed]
猕猴视觉皮层V2到V4和V5区域的通路分离。大自然。1985;315:322 - 325。[PubMed]
猕猴视皮层V5和V1区域之间连接的组织。神经科学杂志1989;1:309-332。[PubMed]
猕猴视皮层V5和V1区域之间连接的组织。神经科学杂志1989;1:333-354。[PubMed]
索茂吉,斯瓦戴,马丁。卡,惠特利奇。猫纹状皮层大篮子细胞形态学和生理特性的突触连接。神经科学。1983;10:261 - 294。[PubMed]
Suzuki W, Saleem KS, Tanaka K.猕猴颞下皮层前部(TE区)的发散向后投影。J Comp Neurol.2000; 422:206 - 228。[PubMed]
Talbot SA, Marshall WH。视觉定位与辨别神经机制的生理学研究。Am J Ophthal。1941;24:125 - 1263。
Thompson JM, Woolsey CN, Talbot SA。兔大脑皮层视觉区I、II。J Neurophysiol。1950;13:277 - 288。[PubMed]
Thompson KG, Hanes DP, Bichot NP, Schall JD。在视觉搜索过程中,猕猴额视野神经元活动的知觉和运动加工阶段被确定。J Neurophysiol。1996;76:4040 - 4055。[PubMed]
Ungerleider LG, Desimone R.从V1和V2的中央和外周场表现投影到颞上沟。J Comp Neurol.1986a; 248:147 - 163。[PubMed]
孙晓燕,张晓燕,张晓燕,等。猕猴视觉区MT的皮层连接。中华神经科学杂志1986b; 248:190-222。[PubMed]
Van Essen DC, Newsome WT, Maunsell JH, Bixby JL。猕猴纹状皮层(V1)到V2和V3区域的投射:不对称、区域边界和斑块连接。中华神经科学杂志1986;244:451-480。[PubMed]
Van Essen DC, Anderson CH Felleman DJ(1992)灵长类视觉系统中的信息处理:一个综合系统的视角。beplay体育公司科学255:419 - 423。
冯·德·海德特,彼得·汉斯,鲍姆加特纳G.错觉轮廓与皮层神经元反应。科学。1984;224:1260 - 1262。[PubMed]
冯·德·海特,彼得·汉斯。猴子视觉皮层的轮廓知觉机制。一、图案不连续线。J > 1989; 9:1731 - 1748。PubMed]
猕猴初级视觉皮层4C层神经元的功能流和局部连接。1998 J >; 18:9489 - 9499。[PubMed]
灵长类动物V1细胞色素氧化酶斑点与2/3层锥体神经元的内在水平连接。Vis Neurosci.1998b; 15:1007 - 1027。[PubMed]
最后更新:2007年6月14日。
作者 马修·斯莫利斯基(Matttew Smolesky)是犹他大学神经生物学和解剖学系的神经科学博士,并获得实验心理学硕士学位。他在研究、教学、行政、联邦政策、指导和拨款写作方面都有成功的记录。他在神经科学和实验心理学方面的经验最初是在视觉知觉和皮质记录领域。在过去的14年里,他一直担任神经科学和心理学项目(韦伯学院,犹他州)的主席或主任。马特在联邦科技政策部门工作了2年。他在合作、项目建设与管理、研究方法与实践、科技交流等方面具有较强的能力。马特目前是美国乔治亚州劳伦斯维尔的乔治亚格温尼特学院的教授和主席。请通过mschmolesky@ggc.edu联系马修·斯莫利斯基 |
 |