Haohua钱gydF4y2Ba
GABA受体的性质gydF4y2Ba
GABA (gydF4y2BaggydF4y2Ba-氨基丁酸)是中枢神经系统的主要抑制性神经递质。GABA的抑制作用是由细胞膜上的受体介导的,并导致神经元兴奋性的降低。至少有三种类型的GABA受体已被鉴定。表1总结了这三种GABA受体的一些一般性质。GABAA受体是配体门控的氯通道。它们介导快速抑制,并广泛分布于整个中枢神经系统。gaba受体具有多种多样的分子组成。6组中至少有16个亚单位已被鉴定。在药理学上,这些受体可被二甲苯拮抗。GABAA受体也是许多治疗化合物(如全身麻醉药、镇静药物和酒精)的靶点。 These compounds allosterically modulate GABAA receptor channel activities. GABAB receptors belong to the G-protein coupled receptor superfamily. The inhibition of GABAB receptors is mediated by indirect gating of either potassium or calcium channels. GABAB receptors are activated by baclofen, and antagonized by phaclofen and saclofen. The subunits of GABAB receptors have recently been cloned. GABAc receptors are the newly identified member of the GABA receptor family. They are also linked to chloride channels, with distinct physiological and pharmacological properties. In contrast to the fast and transient responses elicited from GABAA receptors, GABAc receptors mediate slow and sustained responses. Pharmacologically, GABAc receptors are bicuculline- and baclofen-insensitive, and are not modulated by many GABAA receptor modulators (such as benzodiazepines and barbiturates). GABArgydF4y2Ba亚基被认为参与神经元膜上GABAc受体的形成,但这些受体的确切分子组成尚未确定。gaba acreceptor在大脑多个区域均有表达,主要分布在视网膜神经元上,表明这些受体在视网膜信号处理中起着重要作用。gydF4y2Ba
表1。GABA受体的特征gydF4y2Ba
| 伽马氨基丁酸gydF4y2Ba一个gydF4y2Ba受体gydF4y2Ba | 伽马氨基丁酸gydF4y2BaBgydF4y2Ba受体gydF4y2Ba | 伽马氨基丁酸gydF4y2BaCgydF4y2Ba受体gydF4y2Ba | |
| 类别gydF4y2Ba | Ligand-gated通道gydF4y2Ba | g蛋白耦合的受体gydF4y2Ba | Ligand-gated通道gydF4y2Ba |
| 子单元gydF4y2Ba | A b g d e pgydF4y2Ba | GBR1, GBR2gydF4y2Ba | rgydF4y2Ba |
| 受体激动剂gydF4y2Ba | Muscimol, THIPgydF4y2Ba | 巴氯芬gydF4y2Ba | |
| 拮抗剂gydF4y2Ba | 荷包牡丹碱,苦味毒gydF4y2Ba | PhaclofengydF4y2Ba | TPMPA,苦味毒gydF4y2Ba |
| 脱敏gydF4y2Ba | 是的gydF4y2Ba | 没有gydF4y2Ba | 没有gydF4y2Ba |
| 调制器gydF4y2Ba | 苯二氮平类药物巴比妥酸盐gydF4y2Ba | 锌gydF4y2Ba |
GABAc对视网膜神经元的反应gydF4y2Ba
“GABAc受体”一词最早被Johnston用来描述神经元膜上一个新的GABA结合位点(Johnston, 1986, 1996)。虽然最近的研究表明GABAc受体广泛分布在中枢神经系统的许多部分(Sivilotti和Nistri, 1991;Albrecht等人,1997;Boue-Grabot等人,1998;Wegelius等人,1998;Enz和Cutting, 1999),这些受体在脊椎动物的视网膜中表达最为显著。在白鲈鱼视网膜中,杆状驱动(H4)水平细胞是第一个被鉴定出GABAc受体的视网膜神经元(Qian and Dowling, 1993)。随后,在多种视网膜神经元中检测到gabac受体介导的反应,包括双极细胞(Feigenspan et al., 1993;钱和道林,1995;Qian等,1997b; Lukasiewicz et al., 1994; Lukaisiewicz and Wong, 1997; Pan and Lipton, 1995; Nelson et al., 1999), cone-driven horizontal cells in catfish (Dong et al, 1994; Kaneda et al., 1997), cone photoreceptors (Picaud et al, 1998), and ganglion cells (Zhang and Slaughter, 1995). Among all these retinal neurons, the rod-driven horizontal cells of white perch are the only cells where GABA responses are mediated solely by GABAc receptors. The GABA responses elicited from other cells are usually a mixture of GABA receptors and/or GABA transporters. The unique properties of the white perch rod-driven horizontal cell provided an excellent model to characterize the physiological and pharmacological properties of GABAc receptors on retinal neurons (Qian and Dowling, 1993, 1994).
图1显示了从白鲈鱼视网膜上分离出的一个孤立的杆状水平细胞,以及来自该细胞的典型的gaba诱导反应。这些水平细胞从视网膜的杆状感光细胞接收输入;当分离时,它们在培养中保持典型的形态。杆状水平细胞的胞体是扁平的,胞体直径为50-100 um。有几个粗大的初生枝晶,从中延伸出许多细晶。如图1所示,GABA引起这些细胞缓慢而持续的反应。gaba诱导的膜电流是由氯离子介导的,因此,对这些神经元表现出抑制作用。反应没有脱敏的迹象,即只要GABA存在,反应就保持在一个稳定的水平。此外,这些水平细胞引发的GABA反应表现出缓慢的动力学,这可以在偏移反应中观察到。在GABA应用终止后,膜电流非常缓慢地恢复到基线,时间常数约为15秒。 Such slow and sustained response properties are typical of GABAc receptors.
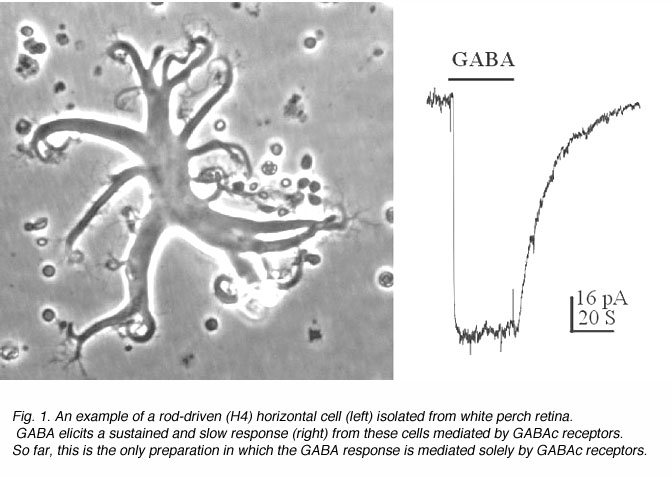
有趣的是,远端视网膜的神经元(即光感受器、水平细胞和双极细胞)根本不产生动作电位。它们对光刺激只能产生缓慢的分级反应。一直以来,人们认为视网膜神经元一定有特殊的方式来处理和分析这种慢信号,而大脑的快速短暂神经元则不然。GABAc的动力学gydF4y2Ba受体介导的反应因此特别适合产生信号的远端视网膜神经元。gydF4y2Ba
GABAc受体的药理学gydF4y2Ba
GABAc受体表现出不同的药理学,不同于传统的GABAA或GABAB受体。GABAc受体由Johnston首次描述为神经元膜上对二甲苯和巴氯芬不敏感的GABA结合位点(Johnston, 1986, 1996)。更详细的研究表明,视网膜神经元上的GABAc受体对GABAA受体(如SR95531和肼)或GABAB受体(如磷氯芬和saclofen)的竞争拮抗剂都不敏感。由于竞争性拮抗剂被认为与受体上的GABA结合位点相互作用,这些结果表明,GABA分子的不同构象是与GABAc受体结合的首选。与这一观点一致的是,GABAA和GABAB受体的特异性激动剂在GABAc受体上表现出截然不同的活性。它们要么没有作用(异吡酯酸、巴氯芬),要么起部分激动剂作用(异胍疫苗、muscimol),要么起拮抗剂作用(THIP、P4S、3-APA和3-APMPA) (Woodward等人,1993;钱和道林,1994)。I4AA是GABAA受体的部分激动剂,是视网膜神经元GABAc反应的有效拮抗剂,也是非洲爪鼠卵母细胞表达GABAc受体的部分激动剂(Qian and Dowling, 1994;Qian等人,1998)。gydF4y2Ba
GABAc受体也不同于经典的GABAA受体,在它们对各种调节器的反应方面。两组化合物,苯二氮杂卓和巴比妥酸盐,是众所周知的调节GABAA活性。另一方面,这些化合物对GABAc受体介导的反应没有任何显著影响(Polenzani et al., 1991)。例如,无论是否使用安定或戊巴比妥(Qian and Dowling, 1993),白鲈鱼杆驱动的水平细胞上的GABA引起的反应几乎是相同的。另一类GABAA受体调节剂,被称为神经活性类固醇,对爪爪卵母细胞中表达的GABAc受体有不同的影响。虽然其中一些可以调节这些受体上的GABA反应,但另一些则不能(Woodward et al., 1992a;Morris等人,1999年)。这些神经活性类固醇对神经元GABAc受体的影响尚未确定。gydF4y2Ba
虽然GABAA和GABAc受体都与氯离子通道相连,但这两种受体的通道性质却有很大的不同。与GABAA受体不同,微辣毒素对GABAc有抑制作用gydF4y2Ba白鲈水平细胞的受体表现出竞争和非竞争机制(Qian and Dowling, 1994)。另一方面,在哺乳动物视网膜(大鼠)中,GABAcgydF4y2Ba受体对微辣毒素阻断不敏感(Feigenspan等人,1993;潘和利普顿,1995)。GABAc的不寻常特征gydF4y2Ba大鼠视网膜上的受体被归因于受体亚基中的单个氨基酸替代(Zhang et al., 1995)。此外,TBPS和丙酸毒素一样是GABA上的另一种氯通道阻滞剂gydF4y2Ba一个gydF4y2Ba受体不会阻断GABAc介导的反应gydF4y2Ba受体(Qian and Dowling, 1994;Woodward等人,1992b)。此外,GABAcgydF4y2Ba受体门控的氯离子通道表现出非常小的单通道电导(Qian and Dowling, 1995;Chang和Weiss, 1999)。gydF4y2Ba
GABAc受体的活性受二价阳离子的调节(Calvo等人,1994;Kaneda等人,1997;Dong和Werblin, 1995;Chang et al., 1995)。特别是,GABAcgydF4y2Ba受体介导的反应被低浓度的锌离子所抑制。GABAc的高灵敏度gydF4y2Ba锌抑制的受体归因于亚基胞外区域的组氨酸残基(Wang et al., 1995)。gydF4y2Ba
最近,一种新的GABAcgydF4y2Ba受体拮抗剂TPMPA已经上市。这种化合物被认为是GABAc的特异性抑制剂gydF4y2Ba受体(Ragozzino et al., 1996)。这种药物的出现将极大地促进这一受体的进一步研究。gydF4y2Ba
GABAc受体的分子生物学gydF4y2Ba
GABAc受体是配体门控通道超家族的成员之一。通过与研究良好的烟碱乙酰胆碱受体类似,GABAc受体被认为表现出如图2所示的结构。这些受体是五聚体,即五个亚基构成功能通道(Amin and Weiss, 1996)。受体有一个长细胞外结构域,包含配体结合位点和几个调节位点。在受体的中间,GABA开启了一个离子通道。GABA与受体的结合引起受体结构的构象变化,从而导致通道的打开。gydF4y2Ba
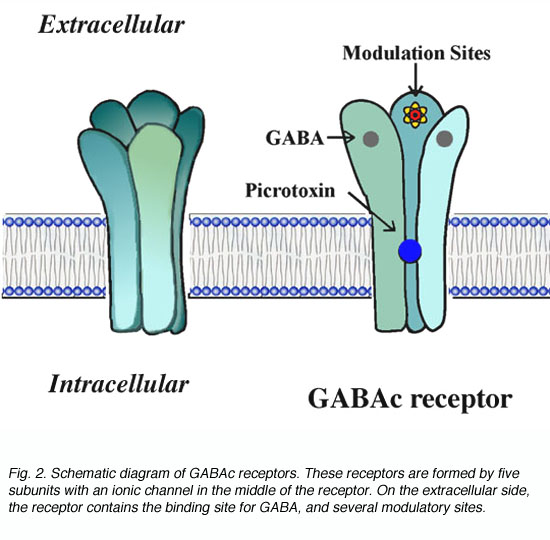
图2所示。GABAc受体示意图。这些受体由五个亚基组成,在受体中间有一个离子通道。在细胞外,受体包含GABA的结合位点和几个调节位点gydF4y2Ba
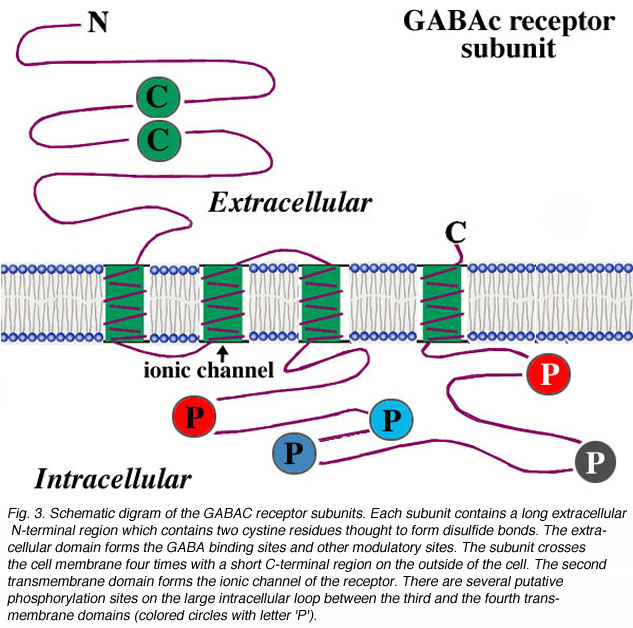
对于形成受体的每个亚基,其结构被认为如图3所示。该亚基包含一个具有配体结合位点的细胞外n端长结构域,四个跨膜结构域,以及一个连接第三和第四个跨膜结构域的细胞内大环。离子通道是由每个亚基的第二个跨膜域形成的。对于细胞内的大环,有几个假定的磷酸化位点,如图中用字母“P”表示。这些残基的磷酸化与受体活性的调节有关。例如,有报道称多巴胺调节鲶鱼锥状水平细胞和虎蝾螈双极细胞末端GABAc受体的活性(Dong和Werblin, 1994;Wellis and Werblin, 1995)。在大鼠双极性细胞上,GABAc受体的活性由蛋白激酶C调节(Feigenspan and Bormann, 1994a)。在GABAc中也观察到胞内第二信使系统对受体活动的调节gydF4y2Ba非洲爪蟾卵母细胞上表达的受体(Kusama et al., 1995)。然而,细胞内第二信使系统的调节机制尚未确定。最近,Filippova和合作者(1999)提供了GABAc的证据gydF4y2Ba亚基磷酸化后的受体内在化。此外,有证据表明GABAc的胞内大环gydF4y2Ba受体亚基参与受体蛋白与其他细胞内蛋白的相互作用,可能在神经元膜上受体聚集起重要作用(Hanley et al., 1999)。gydF4y2Ba
大量的证据表明GABAc受体是由GABA组成的gydF4y2BargydF4y2Ba子单元。这些基因最初是从人类视网膜cDNA文库中克隆而来的(Cutting et al., 1991, 1992)。当在爪蟾卵母细胞中表达时,GABAgydF4y2BargydF4y2Ba亚基形成具有功能的同寡聚受体,其性质与GABAc类似gydF4y2Ba视网膜神经元的受体(Shimada 1992)。此外,GABA的表达gydF4y2BargydF4y2Ba在GABAc所在的视网膜神经元上已检测到亚单位gydF4y2Ba已经记录了受体介导的反应(Qian et al., 1997a;Enz等人,1995,1996)。在白鲈鱼的视网膜上,我们已经克隆了五种形式的GABAgydF4y2BargydF4y2Ba亚单位(Qian et al., 1997a, 1998)。图4显示了白鲈鱼GABA的对比gydF4y2BargydF4y2Ba亚单位和从哺乳动物视网膜克隆的。各种连接元素之间的距离表示亚单元之间的分歧程度。不像哺乳动物的视网膜,只有一种形式gydF4y2BargydF4y2Ba1,gydF4y2BargydF4y2Ba已经确定了2个亚单位,在白栖木中,每个亚单位有两种形式gydF4y2BargydF4y2Ba1,gydF4y2BargydF4y2Ba2家庭。根据推断的氨基酸序列和它们在非洲爪蟾卵母细胞上形成的受体的性质gydF4y2BargydF4y2Ba1,gydF4y2BargydF4y2Ba2家庭被细分为A和B两种形式。全是白栖氨基丁酸gydF4y2BargydF4y2Ba1,gydF4y2BargydF4y2Ba当在爪蟾卵母细胞中表达时,2个亚基能够形成功能性的同源寡聚受体。GABA在这些表达受体中引起的反应是持续的、双环不敏感的,而且不受苯二氮杂卓或巴比妥酸盐的调节,这是GABAc的典型特征gydF4y2Ba受体。像GABAcgydF4y2Ba视网膜神经元上的GABA受体gydF4y2BargydF4y2Ba受体也负责调控氯离子通道。但是,每个GABA的受体都有表达gydF4y2BargydF4y2Ba子单元显示出区别于其他单元的惟一响应属性。例如,GABA活化和微辣毒素抑制的敏感性因亚基而异。此外,I4AA对a型具有拮抗作用gydF4y2BargydF4y2Ba受体,而它是b型的部分激动剂gydF4y2BargydF4y2Ba受体(Qian et al., 1998)。gydF4y2Ba
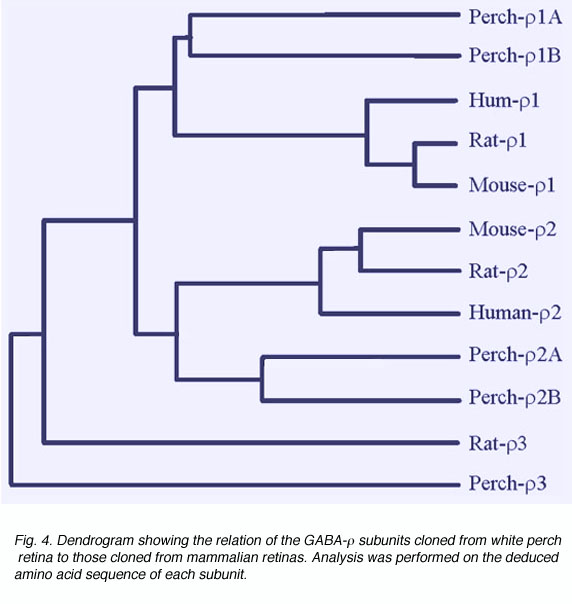
图4。显示GABA-之间关系的树状图gydF4y2BargydF4y2Ba从白鲈鱼视网膜克隆到从哺乳动物视网膜克隆的亚基。对每个亚基推断的氨基酸序列进行分析gydF4y2Ba
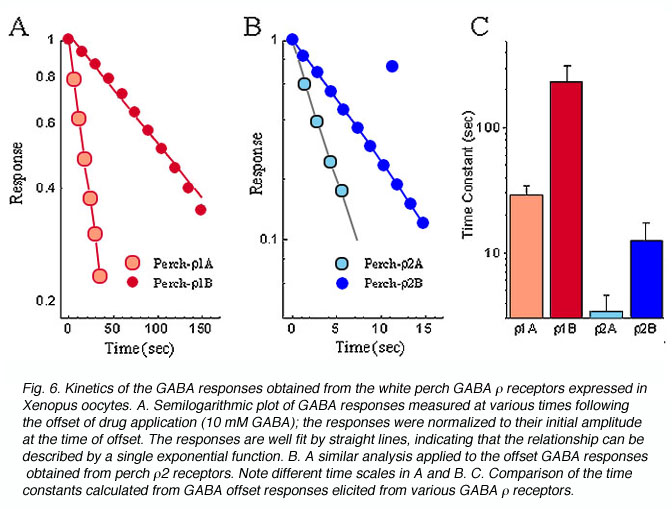
受体介导的反应波形在神经元信号的形成中起着重要的作用。有趣的是,GABA反应的动力学对于每个亚基形成的受体也是不同的。电流跟踪响应应用10gydF4y2Ba米gydF4y2Bamgaba如图5所示。这些记录表明,非洲爪蟾卵母细胞对GABA的反应动力学有显著差异,这取决于哪一种白鲈鱼的GABAgydF4y2BargydF4y2Ba亚基表达。为了量化GABA响应的动力学,偏移GABA响应(终止GABA应用后的电流轨迹)被重新绘制在一个半对数尺度上,振幅归一化到它们的初始值(图6)。在每种情况下,数据都用直线拟合,表明偏移响应可以用单个指数函数来描述。这条线的斜率代表了衰减的时间常数,表明受体所形成的各种gydF4y2BargydF4y2Ba亚单位的反应动力学有显著差异。不同白鲈鱼GABA受体诱导的偏移反应的平均时间常数gydF4y2BargydF4y2Ba亚单位在图6C的柱状图中显示。这两个受体家族及其亚组之间的反应动力学存在一致的差异。例如,来自gydF4y2BargydF4y2Ba1受体明显慢于gydF4y2BargydF4y2Ba2受体。响应动力学的差异gydF4y2BargydF4y2Ba1,gydF4y2BargydF4y2Ba2受体在很大程度上是由亚基的第二个跨膜结构域的单个残基决定的(Qian et al., 1999)。这种二分法中gydF4y2BargydF4y2Ba1,gydF4y2BargydF4y2Ba2个亚基在所有GABA的物种中都很好地保存gydF4y2BargydF4y2Ba亚单位已被克隆。gydF4y2BargydF4y2Ba1个在残基上有脯氨酸的亚基结合形成一个动力学较慢的受体;而gydF4y2BargydF4y2Ba2个亚基在残基处含有丝氨酸,形成反应速度更快的受体。因此,受体构成了人类gydF4y2BargydF4y2Ba1亚基的反应动力学比受体慢gydF4y2BargydF4y2Ba2个亚单位(Enz and Cutting, 1999)。不同GABA形成的受体之间的动力学差异gydF4y2BargydF4y2Ba亚单位可以为神经系统提供构建不同类型的信号过滤器和神经系统中不同类型的神经元信号的基石。gydF4y2Ba
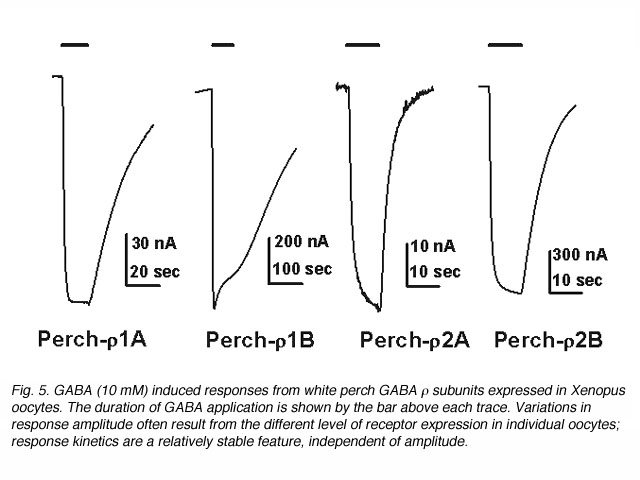
视网膜中GABAc受体的功能gydF4y2Ba
尽管杆状水平细胞提供了一个很好的模型来描述视网膜中的GABAc受体,最近的研究表明GABAc受体也存在于各种其他类型的视网膜神经元上。在鲶鱼的锥形水平细胞中记录到GABAc受体介导的反应(Dong et al., 1994;Kaneda et al., 1997),锥体光感受器(Picaud et al., 1998),以及某些类型的神经节细胞(Zhang and Slaughter, 1995)。迄今为止,在每一种被检测的双极细胞中,GABAc反应都特别显著(Feigenspan等人,1993;钱和道林,1995;Lukasiewicz等人,1994;Lukaisiewicz和Wong, 1997;Qian等人,1997;Nelson等人,1999),免疫细胞化学和原位杂交研究都表明双极细胞上存在GABAc受体(Qian等人,1997;Enz等人,1995,1996; Koulen et al., 1997). It appears that these receptors play an important role in shaping signal transmission from bipolar cells to third order neurons in the retina.
图7说明了从白鲈鱼视网膜分离的双极细胞的一些例子。这些双极性细胞在分离培养时保持其形态。它们通常有一个梨形的细胞体,从细胞体上延伸出几个树突和一个轴突。白鲈鱼视网膜双极细胞的GABA反应既有短暂的成分,也有持续的成分,表明GABAA和GABAc受体都存在,如图8所示。瞬时成分可选择性地通过联合应用双杉碱阻断,留下更持久的反应。因此,双极细胞上GABAc受体的电生理和药理特性与杆状水平细胞上的GABAc受体非常相似(Qian and Dowling 1995;Lukasiewicz等人,1994;Feigenspan和Bormann, 1994)。gydF4y2Ba
 图7所示。培养中单生白鲈鱼双极性细胞的例子gydF4y2Ba |
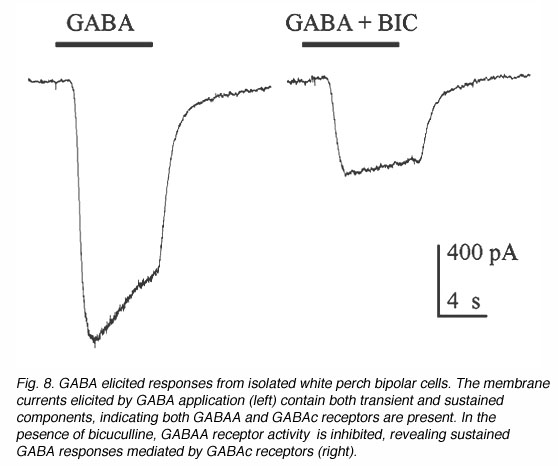 图8所示。GABA引起孤立的白鲈鱼双极性细胞的反应。GABA应用引起的膜电流(左)包含短暂和持续的成分,表明GABAA和GABAc受体都存在。在二甲双胍的存在下,GABAA受体的活性被抑制,揭示了由GABAc受体介导的持续的GABA反应(右)gydF4y2Ba |
GABAA和GABAc受体的不同动力学性质表明它们在介导双极细胞末端抑制方面起着不同的作用(Qian et al., 1997;Lukasiewicz和Shields, 1998)。此外,不同亚型的双极细胞表现出不同比例的GABAA和GABAc受体。例如,在大鼠视网膜中,GABAA和GABAc受体对杆状和锥状双极细胞的贡献存在明显差异(Euler和Wassle, 1998)。在白鲈鱼中,不同形态类型的双极细胞表现出不同比例的GABAc受体介导成分(Qian and Dowling, 1995)。这些结果强烈表明,不同亚型的双极细胞利用各种GABAA和GABAc受体的混合物执行不同的活动,并帮助创建通过视网膜的各种功能通路。gydF4y2Ba
由于视网膜神经元上存在多个GABA受体,有时很难分离每个受体的贡献。最近对神经节细胞反应的研究揭示了GABAc受体在视网膜信息处理中的一些有趣特征。例如,GABAc受体的激活导致神经节细胞产生更多的瞬变光反应(Dong和Werblin, 1998), GABAc受体介导的延迟抑制被认为在神经节细胞感受野的边缘增强塑造中起着重要作用(Jacobs和Werblin, 1998)。双极细胞到神经节细胞突触可能受抑制性大分泌前馈或反馈突触的严重影响,这些似乎主要是通过GABAc受体。gydF4y2Ba
参考文献gydF4y2Ba
Albrecht BE, Breitenbach U, Stuhmer T, Harvey RJ, Darlison MG。原位杂交和逆转录聚合酶链反应研究GABA(C)受体rho1和rho2亚基基因在禽和大鼠脑中的表达。中华神经科学杂志1997;9:2414-2422。[gydF4y2BaPubMedgydF4y2Ba]gydF4y2Ba
Amin J, Weiss DS。通过野生型和激活受损亚基的共同表达获得rho1 GABA受体激活机制的研究。中国生物医学工程学报1996;23(3):393 - 393。[gydF4y2BaPubMedgydF4y2Ba]gydF4y2Ba
张志强,张志强,张志强,等。GABA受体rho亚基在大鼠脑组织中的表达。J Neurochem。1998;70:899 - 907。[gydF4y2BaPubMedgydF4y2Ba]gydF4y2Ba
张海燕,王晓燕,张晓燕,等。非洲爪蟾卵母细胞中r1型g-氨基丁酸受体的阳离子调控。中国科学:地球科学(英文版)1994;91:12725-12729。[gydF4y2BaPubMedgydF4y2Ba] [gydF4y2Ba免费全文PMCgydF4y2Ba]gydF4y2Ba
Chang Y, Amin J, Weiss DS。锌是同聚体r1 g-氨基丁酸活化通道的混合拮抗剂。摩尔杂志。1995;47:595 - 602。gydF4y2BaPubMedgydF4y2Ba]gydF4y2Ba
Chang Y, Weiss DS。通道打开将激动剂锁定在GABAC受体上。Nat > 1999; 2:219 - 225。[gydF4y2BaPubMedgydF4y2Ba]gydF4y2Ba
切割GR、Lu L、Zoghbi H、O’hara BF、Kasch LM、monrose - rafizader C、Donovan DM、Shimada S、Antonarakis SE、Guggino W、Uhl GR、Kazazian HH。g-氨基丁酸(GABA) rho1 cDNA的克隆:视网膜高表达的GABA受体亚基。中国科学(英文版)1991;88:2673-2677。[gydF4y2BaPubMedgydF4y2Ba] [gydF4y2Ba免费全文PMCgydF4y2Ba]gydF4y2Ba
剪切GR, Curristin S, Zoghbi H, O’hara B, Selden MF, Uhl GR.确定γ -氨基丁酸(GABA)受体亚基rho2 cDNA,并将编码rho2 (GABRR2)和rho1 (GABRR1)的基因与人6q14-q21和小鼠4号染色体共定位。Genomics.1992; 12:801 - 806。[gydF4y2BaPubMedgydF4y2Ba]gydF4y2Ba
Dong CJ, Picaud SA, Werblin FS。鲶鱼锥体上的GABA转运体和gabac样受体——但不是杆状驱动的水平细胞。J Neurosci.1994; 14:2648 - 2658。[gydF4y2BaPubMedgydF4y2Ba]gydF4y2Ba
Dong CJ, Werblin FS。多巴胺对离体视网膜神经元GABAC受体功能的调节。J Neurophysiol。1994;71:1258 - 1260。[gydF4y2BaPubMedgydF4y2Ba]gydF4y2Ba
Dong CJ, Werblin FS。锌下调调节从鲶鱼视网膜上分离出来的锥形水平细胞的GABAc受体电流。J Neurophysiol.1995; 73:916 - 919。[gydF4y2BaPubMedgydF4y2Ba]gydF4y2Ba
Dong CJ, Werblin FS。通过GABAC反馈的时间对比增强在双极终端在老虎蝾螈视网膜。J Neurophysiol.1998; 79:2171 - 2180。[gydF4y2BaPubMedgydF4y2Ba]gydF4y2Ba
Enz R, Brandstätter JH, Hartveit E, Wässle H, Bormann J. GABA受体rho 1和rho2亚基在大鼠视网膜和大脑中的表达。中华神经科学杂志1995;7:1495-1501。[gydF4y2BaPubMedgydF4y2Ba]gydF4y2Ba
张志强,张志强,张志强,等。哺乳动物视网膜GABAC受体rho亚基的免疫细胞化学定位。J Neurosci.1996; 16:4479 - 4490。[gydF4y2BaPubMedgydF4y2Ba]gydF4y2Ba
GABAC受体rho亚基在人中枢神经系统中异质表达,形成具有不同物理性质的均聚物和异聚物。中华神经科学杂志1999;11:41-50。[gydF4y2BaPubMedgydF4y2Ba]gydF4y2Ba
GABAA和GABAC受体在大鼠视网膜切片制备中对棒和锥双极细胞的不同贡献。J Neurophysiol.1998; 79:1384 - 1395。[gydF4y2BaPubMedgydF4y2Ba]gydF4y2Ba
张志强,张志强,张志强,等。GABA受体的药理作用gydF4y2Ba- - - - - -gydF4y2Ba大鼠视网膜双极细胞中的通道。大自然。1993;361:159 - 163。[gydF4y2BaPubMedgydF4y2Ba]gydF4y2Ba
张志强,张志强。GABAC受体对视网膜双极细胞GABAC受体的调控作用。中国生物医学工程学报。1994,19(3):323 - 326。[gydF4y2BaPubMedgydF4y2Ba] [gydF4y2Ba免费全文PMCgydF4y2Ba]gydF4y2Ba
张志强,张志强。GABAA和GABAC受体对大鼠视网膜双极细胞的差异药理作用。药理学进展,1994b; 288:97-104 .[gydF4y2BaPubMedgydF4y2Ba]gydF4y2Ba
Filippova N, Dudley R, Weiss DS。重组人r1 GABAC受体磷酸化依赖内化的证据。J Physiol.1999; 518:385 - 399。[gydF4y2BaPubMedgydF4y2Ba]gydF4y2Ba
Hanley JG, Koulen P, Bedford F, Gordon-Weeks PR, Moss SJ。蛋白质MAP-1B连接GABA(C)受体到视网膜突触的细胞骨架。1999; 397:66 - 69。[gydF4y2BaPubMedgydF4y2Ba]gydF4y2Ba
Jacobs AL, Werblin FS。视网膜输出的时空模式。J Neurophysiol。1998;80:447 - 451。[gydF4y2BaPubMedgydF4y2Ba]gydF4y2Ba
约翰斯顿雀鳝。GABA受体的多样性。图片来源:Olsen RW, Venter JC,编辑。受体生物化学与方法学。卷。5。Alan R. Liss, Inc;1986.57 - 71页。gydF4y2Ba
约翰斯顿雀鳝。GABAc受体:相对简单的发射门控离子通道?中国生物医学工程学报1996;21(5):319 - 323。[gydF4y2BaPubMedgydF4y2Ba]gydF4y2Ba
Kaneda M, Mochizuki M, Kaneko A. catfish视网膜水平细胞中GABAC反应的Ca2+和其他二价阳离子调控。中华物理杂志1997;110:741-747。[gydF4y2BaPubMedgydF4y2Ba]gydF4y2Ba
Koulen P, Brandstätter JH, Kröger S, Enz R, Bormann J, Wässle H. GABA(C)受体rho亚基在猫、金鱼和鸡视网膜中的免疫细胞化学定位。中华神经科学杂志1997;380:520-532。[gydF4y2BaPubMedgydF4y2Ba]gydF4y2Ba
草间敏,刘志刚,刘志刚,村上春树。GABA rho1受体:蛋白激酶C激活物的抑制作用。中华药理学杂志1995;29(3):431 - 434。[gydF4y2BaPubMedgydF4y2Ba]gydF4y2Ba
Lukasiewicz PD, Maple BR, Werblin FS。蝾螈视网膜双极细胞末端的GABA受体。J > 1994; 14:1202 - 1212。gydF4y2BaPubMedgydF4y2Ba]gydF4y2Ba
Lukasiewicz警署,Wong ROL。雪貂视网膜双极细胞上的GABAC受体:哺乳动物亚型的多样性?Vis > 1997; 14:989 - 994。gydF4y2BaPubMedgydF4y2Ba]gydF4y2Ba
GABAA和GABAC受体的不同组合赋予视网膜突触反应不同的时间特性。J Neurophysiol。1998;79:3157 - 3167。[gydF4y2BaPubMedgydF4y2Ba]gydF4y2Ba
神经活性类固醇对γ -氨基丁酸C型受体的微分调节。摩尔Pharmacol.1999; 56:752 - 759。[gydF4y2BaPubMedgydF4y2Ba]gydF4y2Ba
李玉霞,李志强,李志强。GABA(C)样反应在大鼠视网膜神经细胞中的分布。Vis Neurosci.1999; 16:179 - 190。[gydF4y2BaPubMedgydF4y2Ba]gydF4y2Ba
潘Z-H,立顿萨。多种GABA受体亚型介导大鼠视网膜双极细胞末梢钙内流抑制。J > 1995; 15:2668 - 2679。[gydF4y2BaPubMedgydF4y2Ba]gydF4y2Ba
黄晓明,张晓明,张晓明,张晓明。GABAA和GABAC受体在成年猪锥细胞中的表达:来自光受体-神经胶质细胞共培养模型的证据。杂志。1998;513:33-42。[gydF4y2BaPubMedgydF4y2Ba]gydF4y2Ba
张志强,张志强,张志强,等。哺乳动物g-氨基丁酸受体在非洲爪蟾卵母细胞中的表达。中国科学(英文版)1991;88:4318-4322。[gydF4y2BaPubMedgydF4y2Ba] [gydF4y2Ba免费全文PMCgydF4y2Ba]gydF4y2Ba
H倩,王道林。杆状视网膜水平细胞对GABA的新反应。大自然。1993;361:162 - 164。[gydF4y2BaPubMedgydF4y2Ba]gydF4y2Ba
H倩,王道林。白鲈鱼视网膜杆水平细胞上发现的新型GABA受体的药理学研究。J > 1994; 14:4299 - 4307。gydF4y2BaPubMedgydF4y2Ba]gydF4y2Ba
H倩,王道林。杂合低音视网膜双极细胞GABAA和GABAC受体。J Neurophysiol。1995;74:1920 - 1928。[gydF4y2BaPubMedgydF4y2Ba]gydF4y2Ba
H钱,Hyatt G, Schanzer A, Hazra R, Hackam A, Cutting GR, Dowling JE。白鲈鱼视网膜GABAC和rho亚基受体的比较。Vis > 1997; 14:843 - 851。[gydF4y2BaPubMedgydF4y2Ba]gydF4y2Ba
李丽娟,李丽娟,张晓燕,张晓燕,张晓燕。双极细胞GABA受体的研究进展:锌对GABA介导膜电流的作用。J Neurophysiol。1997 b; 78:2402 - 2412。[gydF4y2BaPubMedgydF4y2Ba]gydF4y2Ba
刘建平,张建平,刘建平。白鲈鱼视网膜中GABA-rho亚基的分子生物学特性。J一般。1998;37:305 - 320。gydF4y2BaPubMedgydF4y2Ba]gydF4y2Ba
Qian H, Dowling JE, Ripps H. GABA r亚基第二跨膜结构域的单个氨基酸是GABAC受体反应动力学的决定因素。J一般。1999;40:67 - 76。[gydF4y2BaPubMedgydF4y2Ba]gydF4y2Ba
Ragozzino D, Woodward RM, Murata Y, Eusebi F, Overman LE, Miledi R.选择性γ -氨基丁酸C受体拮抗剂的设计和体外药理学。摩尔杂志。1996;50:1024 - 1030。[gydF4y2BaPubMedgydF4y2Ba]gydF4y2Ba
岛田S, Cutting G, Uhl GR. G -氨基丁酸A或C受体?g-氨基丁酸r1受体RNA诱导非洲爪蟾卵母细胞中双环碱-、巴比妥酸-和苯并二氮杂卓-不敏感的g-氨基丁酸反应。摩尔杂志。1992;41:683 - 687。[gydF4y2BaPubMedgydF4y2Ba]gydF4y2Ba
GABA受体在中枢神经系统中的作用机制。食物一般。1991;36:35 - 92。[gydF4y2BaPubMedgydF4y2Ba]gydF4y2Ba
单组氨酸残基是锌抑制GABA rho 1受体所必需的。J Neurosci.1995; 15:7684 - 7691。[gydF4y2BaPubMedgydF4y2Ba]gydF4y2Ba
Wegelius K, Pasternack M, Hiltunen JO, Rivera C, Kaila K, Saarma M, Reeben M. GABA受体rho亚基转录本在大鼠脑中的分布。中华神经科学杂志1998;10:35 - 357。[gydF4y2BaPubMedgydF4y2Ba]gydF4y2Ba
Wellis DP, Werblin FS。多巴胺调节GABAc受体介导抑制钙进入双极细胞终端和递质释放在虎蝾螈视网膜。J > 1995; 15:4748 - 4761。[gydF4y2BaPubMedgydF4y2Ba]gydF4y2Ba
黄晓明,李晓峰,李晓峰,等。激素对哺乳动物脑和视网膜poly(A)+ RNA表达g-氨基丁酸受体的影响。摩尔杂志。1992;41:89 - 103。[gydF4y2BaPubMedgydF4y2Ba]gydF4y2Ba
刘文华,刘文华,刘文华,等。非洲爪蟾卵母细胞中对二甲苯/巴氯芬不敏感的g-氨基丁酸受体的表达研究。摩尔杂志。1992;42:165 - 173。[gydF4y2BaPubMedgydF4y2Ba]gydF4y2Ba
作者:刘文华,刘文华,刘文华,等。非洲爪蟾卵母细胞中表达的双单克林/巴氯芬不敏感(r-like) g-氨基丁酸受体的研究。2g-氨基丁酸A和g-氨基丁酸B受体激动剂和拮抗剂的药理学研究。摩尔杂志。1993;43:609 - 625。gydF4y2BaPubMedgydF4y2Ba]gydF4y2Ba
张迪,潘志华,张欣,AD新娘,立顿萨。大鼠视网膜γ -氨基丁酸C型受体亚基的克隆与通道阻塞中微量毒素的蛋氨酸残基。中国科学:地球科学(英文版)1995;[gydF4y2BaPubMedgydF4y2Ba] [gydF4y2Ba免费全文PMCgydF4y2Ba]gydF4y2Ba
GABAC受体对两栖动物视网膜ON通路的选择性抑制。J Neurophysiol。1995;74:1583 - 1592。gydF4y2BaPubMedgydF4y2Ba]gydF4y2Ba
| 作者gydF4y2Ba划归博士昊华钱gydF4y2Ba出生于中国江苏。1982年在南京大学获得生物学学士学位,1985年在上海生理研究所获得神经生物学硕士学位,1991年在芝加哥伊利诺伊大学获得解剖学和细胞生物学博士学位。在哈佛大学与John E. Dowling博士进行博士后研究期间,他描述了一种新的GABA受体,即视网膜神经元上的GABAc受体。1996年,他加入伊利诺伊大学芝加哥分校眼科和视觉科学系,继续研究脊椎动物视网膜中GABAc受体的分子结构和生理功能。2010年,他转至美国国家眼科研究所,目前担任该研究所所长gydF4y2Ba视觉功能核心设施gydF4y2Ba.核心已经开发了许多新的方法来评估各种视网膜疾病动物模型的视觉功能。gydF4y2Ba |
