Michael Kalloniatis和Charles Luu
概述。
灵长类视觉系统的颜色视觉处理是由三种不同光谱类别的视锥细胞对光的吸收启动的。beplay体育公司因此,色彩视觉被描述为三变色或三色,最初的心理物理学研究表明,颜色可以通过使用三种不同的三原色来匹配。1802年,托马斯·杨提出了一个模型,认为颜色的感知可以由三个主要的颜色感受器编码,而不是由数千个颜色感受器编码单个颜色。
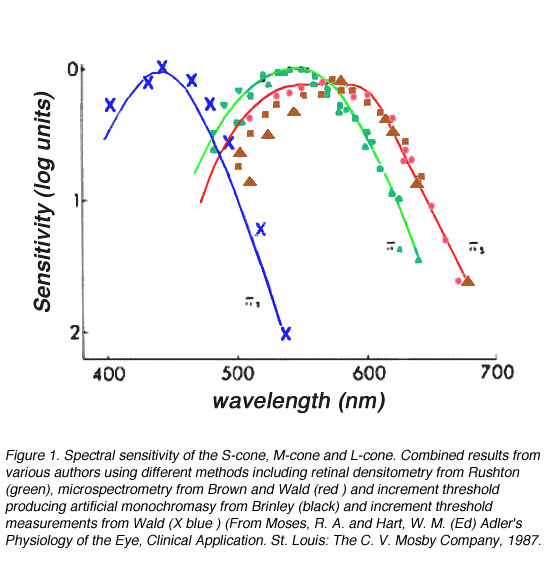
锥的光谱灵敏度可以通过几种方法来确定。其中两种方法包括分离受体反应(Baylor等人,1984年),利用法线和二色体的颜色匹配函数进行计算(Smith和Pokorny, 1975年;重色谱仪是指视网膜上有一个锥体光颜料缺失的对象)、微光谱法(Bowmaker和Dartnall, 1980)或反射密度法(Rushton, 1963, 1966)。微能谱仪技术包括隔离单个锥体并让光通过它。不同波长的透射率变化可以用来计算锥的光谱吸收或确定电响应的变化。反射密度测定法涉及到在视网膜中引导光线,并确定吸收的变化作为波长的函数。这些结果随后被用来计算光谱吸收。
从上述技术中已分离出人类视网膜中的三类视锥细胞。这三类视锥分别是短波敏感(s -cone)、中波长敏感(m -cone)和长波长敏感(l -cone),它们的光谱灵敏度不同但重叠。校正视网膜前光损失后,s -cone的光谱灵敏度在大约440 nm处峰值,m -cone的峰值在545 nm处,l -cone的光谱灵敏度在565 nm处峰值,尽管不同的测量技术导致的最大灵敏度值略有不同(图1)。
颜色匹配
色觉的三色特性使几乎任何颜色都能与三种颜色的混合相匹配。这种三色视觉也是线性的。这意味着比色方程具有普通方程的性质。
颜色与数量相配C年代一个刺激的年代可以表示为:
Cs(S) = C1(l1) + C2(l2) + C3(l3)
在哪里李三原色(不一定是单色光谱色)和Ci三色值。
三刺激值表示实现颜色和光度匹配所需的三个基色中的每个基色的数量(图2)。它们可以用光通量或辐射通量的单位表示,甚至可以用任意刻度表示,前提是任意刻度表示这三个量的相对比例。

图2。受试者被要求调整二部场左侧的三种颜色,以匹配二部场右侧的标准颜色
加色和减色混合
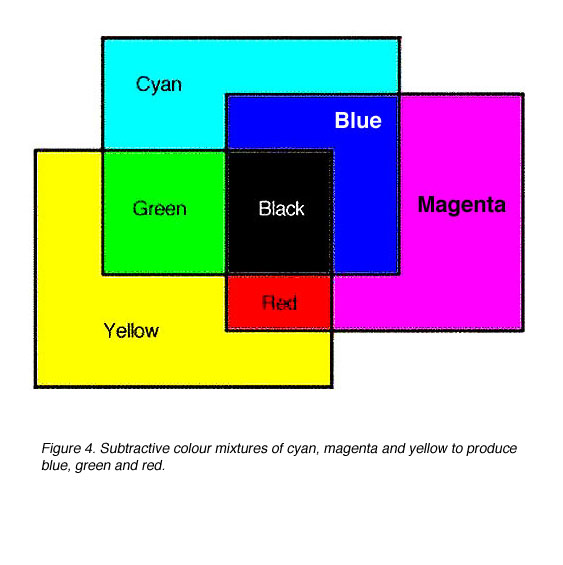
两种或两种以上的颜色可以混合在一起产生一种由最初使用的颜色混合而成的新颜色。这可以在白色屏幕上演示,有三种颜色:蓝色、绿色和红色。青色、品红、黄色和白色是由这些颜色混合而成的(图3)。在选择颜色混合实验中使用的颜色时,要求之一是其中两种颜色不能混合产生第三种颜色。

减色法混合颜色涉及对波长的选择性吸收。青色、品红和黄色是减色法原色。如果白光通过黄色滤光片照射,黄色滤光片就会吸收蓝色,透射红色和绿色(从而产生黄色)。因此,黄色可以被认为是-B滤波器。洋红色滤光片从白光中减去或吸收绿色(-G滤光片),青色滤光片从白光中减去或吸收红色(-R滤光片)(图4)。
Grassmann定律提供颜色匹配数据的定量描述(格拉斯曼1855)。它们在规定的一系列条件下(关于亮度、观察者的适应性、场的大小等)都能保持良好。格拉西曼定律在量化配色数据时很有用,但作为一种规则,配色数据受以下因素的影响:
- 受试者中央4 - 5度视力的黄斑色素(叶黄素)变化
- 色差
- 棒侵入,特别是当大场和低光水平涉及
- 亮度可加性的阿布尼定律失效,包括亥姆霍兹-科尔劳什效应
- Bezold-Brücke明亮强度时的色相变化
颜色规范。
孟塞尔色彩系统
所有颜色都可以根据它们的色调、明度和饱和度来完全指定。蒙塞尔系统有三个维度:色相、值和色度。这三个维度对应着人类色彩视觉的三个感知属性。

蒙塞尔色彩系统的三个维度是:
1.色调:有关波长或主波长.色相是由字母和数字组成的100步刻度的组合表示的(图5)。有十个字母类别用于表示色相,每个字母类别进一步细分(通过使用数字1到10)为十个子组。如果表示色调子组的数字是5,那么它可以被省略。5R和R的色调相同)。
2.值:值以从1(黑色)到10(白色)的数值刻度指定,该属性与反射而且光度(或亮度)。
3.浓度:色度是蒙塞尔项对应的饱和.它的数值范围为0到各种最大值,取决于可用猪液可获得的饱和度。
例如,一种颜色的记号可能是2GY 6/10。这意味着它是一种非常接近黄色的绿色/黄色;它的值是6。几乎在黑白比例的中间)和10的色度(即。饱和)。
孟塞尔系统中使用的缩放被设计成感知上一致的。换句话说,颜色样本按相同的视觉步骤排列。例如,色度3和色度4之间的感知差异(名义上)与色度4和色度5之间的感知差异相同。尽管不同维度上的步长不同,但这种缩放对于所有三个维度都是相同的不可比性(也就是说,色调上的单步差异与饱和度上的单步差异不具有相同的感知差异)。
蒙塞尔系统的示意图如图6所示。数值刻度在垂直轴上,色调刻度在圆柱体的周长上,色度刻度在径向刻度上。图5所示的Munsell陀螺是这个概念的更精确的表示。

1931年国际色度委员会(CIE)色度系统
使用三种颜色的三色配色可以在NewtonÕs颜色圈上说明(图7)。牛顿的颜色圈由以下组件组成:
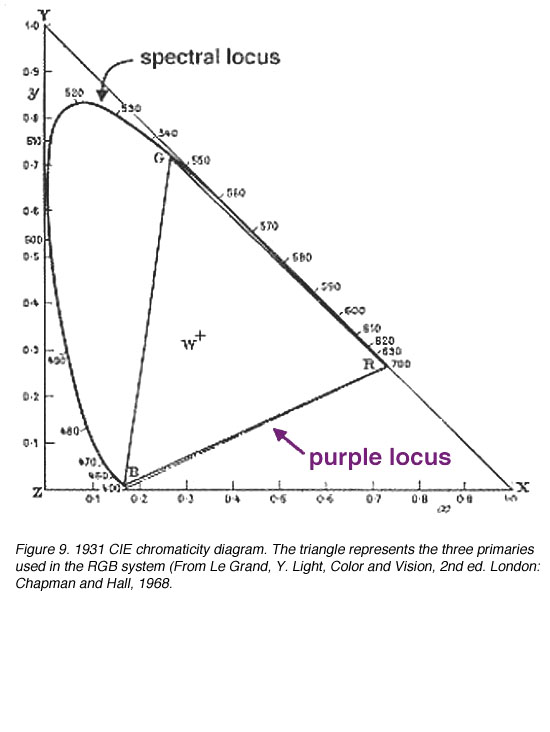
- 代表光谱颜色的圆圈(虽然这里没有显示,但蓝色和红色的混合物(紫色轨迹不是光谱颜色,因此R和B之间的直线更合适)
- 一个三角形,它的顶点代表用于颜色匹配的三种原色(R, G和B)
- 圆心代表白色(W)

牛顿的颜色圈提供了颜色匹配的定性描述,可以用来解释为什么两种颜色可能不足以使颜色匹配,以及使用“负”颜色。例如,如果需要匹配500纳米(位于圆上的光谱颜色),则需要蓝色和绿色。然而,单独使用蓝色和绿色原色会产生500纳米的去饱和光。因此,必须在光谱颜色中加入红色来降低它的饱和度并进行匹配(图8)。当加入第三个原色来降低混合颜色的饱和度时,会得到负的三刺激值(图5)。这可以用下面的等式来证明:
500 nm (S) + RºG + B
或
500海里(S)ºG + B- R

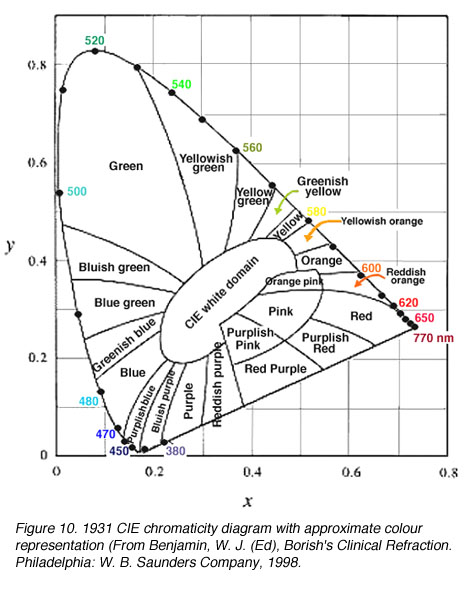
为了处理“负”色,CIE设计了XYZ系统,使用非真实的(想象的)三原色来描述颜色空间。1931 CIE色度系统选择了三个假想的初级(参考刺激)X, Y和Z,这样所有位于这个三角形内的光谱位点都是正的。alychne是没有亮度的颜色轨迹,它被选在XYZ色度系统的X到Z上。所有的光度都用Y表示,选择Y的参考位点只是为了包围真实颜色的区域。选择等能量的白色具有相同的色度坐标,即0.33,0.33(图9和10)。色度坐标表示三个原色的相对贡献,坐标之和为1.0。因此,z可以通过知道坐标x和y来计算,因为x + y + z = 1。
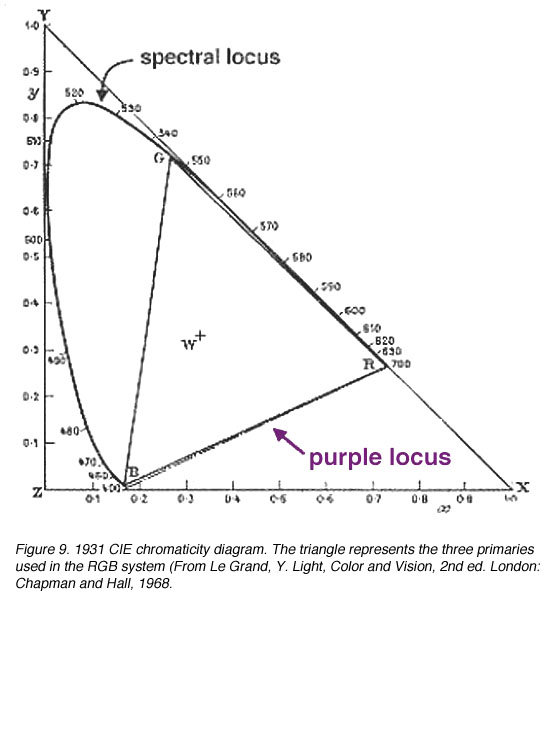 图9。1931 CIE色度图。三角形代表在RGB系统中使用的三种主要(来自Le Grand, Y.光,颜色和视觉,2nd伦敦:查普曼和霍尔出版社,1968年 |
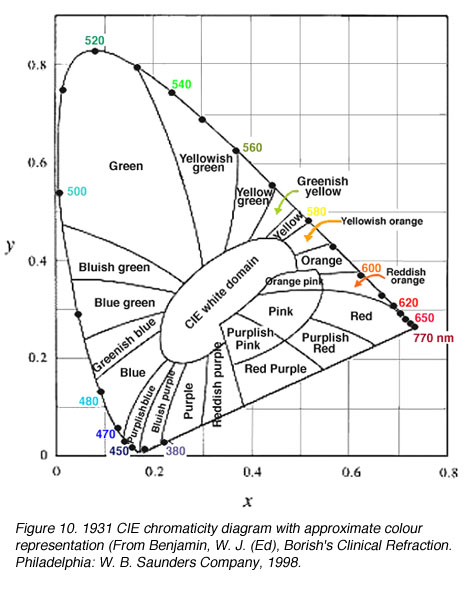 图10。1931 CIE色度图的近似颜色表示(来自Benjamin, W. J. (Ed), Borish的临床折射。费城:W. B.桑德斯公司,1998 |
{kind=link}
{kind=link}
样品的主导波长、补充波长和激发纯度很容易确定。主导波长代表颜色的主要波长。互补波长是在与主导波长混合适当部分时产生白色的波长。当一条线与另一条经过消色差点(如图11所示)的线相连时,可以发现谱补。A的主导波长由DA处的光谱波长给出,由CA处的光谱波长补充(图11)。C点是白点的位置,B点是另一个波长,当以适当的比例混合时,会产生白色。

图11。颜色A的互补和主要光谱波长,颜色B也是颜色A的互补,因为两种波长的适当混合会产生白色
消色差点的变化取决于所使用的标准光源(图12)。一种转变x而且y坐标发生在色温升高的时候。对于标准光源C,绿光没有互补的波长(波长在492 nm到567 nm之间)。然而,适当选择的紫光可以形成白光(图12)。

图12。消色差点随色温变化的位置。(选自Benjamin, W. J. (Ed), Borish的临床折射。费城:W. B.桑德斯公司,1998)
辨色力函数。
色相、饱和度和亮度这三个颜色视觉变量都取决于波长。辨色实验让我们知道,要检测到色相、饱和度和亮度的差异,波长需要变化多少。
色调的歧视描述波长的变化量(l +DL),这需要能够检测到色调的变化。对于蓝光和红光,要检测到色相的变化,波长需要有很大的变化,而对于具有正常色觉的人来说,大部分光谱的波长变化不超过2纳米(图13)。

图13。平均波长分辨曲线。(摘自戴维森,《眼睛》第二卷。伦敦,文献出版社,1962年)
饱和的歧视描述颜色的苍白程度。饱和度与比色纯度(P)有关,比色纯度也被定义为:
P = L / (Lw + L)在哪里l是光谱的亮度和颜色Lw是混合了光谱色的白色的亮度。一种颜色的比色纯度量化了与光谱色混合的白色的数量。如果光谱颜色是纯的(没有添加白色),则比色纯度为1。
在饱和度判别实验中,亮度保持恒定。双面场使用白色(Lw)在一侧,白色与光谱颜色混合在另一侧(Lw +)DL).在图14中可以看到,需要更多的570纳米颜色才能使白色贴片呈现彩色。因此,黄色的饱和功率较低,而蓝色和红色的饱和功率较高。

图14。Priest和Brickwide(1938)的饱和度辨别来自Graham, c.h., (ed),《视觉与视觉知觉》。纽约:John Wiley and Sons, Inc., 1965)
V (l)功能与亮度的感觉非常匹配,因此通常被认为是反射亮度的歧视.波长555 nm被认为是光谱中最亮的(见前面关于光和暗光度函数的部分)。
色彩视觉理论。
任何色彩视觉理论都必须预测前面提到的所有感知属性。我们在这里提出了一个简单的观点,三色和颜色的对立理论。参见Peter Gouras的“色彩视觉”部分,了解颜色对抗的生理学相关的讨论。
的三色理论1802年由托马斯·杨首次提出,1866年由赫姆霍尔兹进一步探索。这一理论主要基于色彩混合实验,并提出三种通道的组合可以解释色彩识别功能。
三色理论的证据包括:
- 用Rushton视网膜密度法测定两种锥状色素的光谱灵敏度(Rushton, 1963)。
- 用微光谱法鉴定三种锥状颜料(Marks, Dobelle和MacNichol, 1964)。
- L、M和S球果遗传密码的鉴定(nathan et al, 1986a,b)。
- 颜色匹配函数。
- 分离光感受器并测量其作为波长函数的生理反应(Baylor et al, 1984)。
- 光谱灵敏度测量(Wald-Marre光谱灵敏度函数和Stiles光谱灵敏度函数)p机制)
然而,分色理论未能解释红、绿、黄、蓝这四种独特的颜色,也未能解释为什么二色者能感知白色和黄色。它也未能充分解释颜色辨别功能和对手颜色感知。
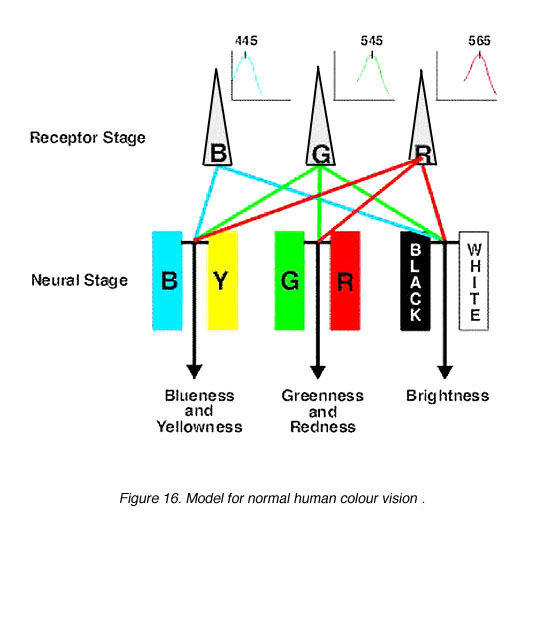
的对手色彩理论由海林在1872年首次提出。当时,这一理论与广为接受的三色理论相抗衡,三色理论解释了视觉的三色并预测了颜色的匹配。海林的对抗性色理论认为有三种通道:红绿、蓝黄和黑白,每一种通道都以对抗性的方式反应。也就是说,只有红色或绿色被感知,而没有绿红色。然而,海林从未挑战三色理论所表达的加工的初始阶段。他只是认为,任何色彩视觉理论都应该解释我们的知觉,也就是彩色图像所揭示的色彩对立。
Hurvich和Jameson(1957)提供了颜色对立的量化数据。利用色相抵消范式,分离出心理物理色相对立通道。Vl用亮度判别函数描述对黑白的感知。因此,通过调节蓝色或黄色和红色或绿色的数量,可以匹配任何样本波长(图15)。除了四种独特的色调(蓝、绿、黄、红)外,所有波长的互补波长都可以相互抵消。

支持对手颜色理论的其他证据包括:
- 鱼视网膜水平细胞的电记录显示蓝黄色的对手过程和红绿色的对手(Svaetichin, 1956)。
- 侧膝状核的电记录显示相反的颜色过程(DeValois et al, 1966)。
- 灵长类动物视网膜神经节细胞的电记录显示相反的颜色过程(Gouras, 1968;de Monasterio和Gouras, 1975;Zrenner和Gouras, 1981)。
阶段理论:这导致了正常色彩视觉的现代模型,该模型将三色理论和对手色彩理论分为两个阶段(图16)。第一阶段可以被认为是受体阶段,由三种光色素(蓝、绿、红锥)组成。第二个是神经处理阶段,即颜色对立发生的阶段。第二阶段发生在受体后水平,最早发生在水平细胞水平。
彩色视觉缺陷。
色盲可能是先天的,也可能是后天的。先天性色觉缺陷(CVD)在出生时就存在,是遗传的,而获得性色觉缺陷是继发于眼病。红绿色觉障碍和蓝锥单色症以x连锁隐性模式遗传。OPN1LW(长波长)和OPN1MW(中波长)基因位于X染色体上。在男性(只有一条X染色体)中,每个细胞中的一个基因变化就足以引起这种情况。男性受X连锁隐性遗传病影响的频率(8%)要比女性(0.5%)高得多(表1),因为女性(有两条X染色体)必须在染色体的两个拷贝上都发生遗传变化才能引起这种疾病。x连锁遗传的一个特征是父亲不能将x连锁遗传给儿子。
蓝黄色视觉缺陷是一种常染色体显性遗传模式,这意味着每个细胞中一个被改变的OPN1SW(短波长)基因拷贝就足以引起这种情况。在许多情况下,受影响的人从受影响的父母那里继承了这种情况。(来自ghr网站,nim.nih.gov遗传学家参考“色觉缺陷”。国家医学图书馆;狄(2004))。甚至有证据表明,色盲患者视网膜的锥体结构发生了改变(Patterson et al. 2016)。
心血管疾病分为三类。它们是单色、二色和异常三色。色觉正常的人被称为三色视者。单色者通常是完全色盲的,除了杆状通路外可能还有一个锥状通路。二色体缺少一个锥体光色素,因此,它们只有两个锥体通道。异常三种锥状光颜料均为异常三种锥状光颜料,但其中一种锥状光颜料为异常,其峰值灵敏度移位。
表1先天性颜色缺陷患病率
| 色彩视觉的类型缺乏 | 男性 | 女性 |
| 整体 | ~ 8% | ~ 0.5% |
| 异常trichromasy | ||
| protanomaly | 1% | 0.01% |
| deutanomaly | 5% | 0.4% |
| tritanomaly | 罕见的 | 罕见的 |
| Dichromasy | ||
| 红色盲 | 1% | 0.01% |
| 绿色盲 | 1.5% | 0.01% |
| 蓝色盲* | 0.008% | 0.008% |
| 全色盲 | ||
| 杆全色盲 | 罕见的 | 罕见的 |
| 锥全色盲 | 罕见的 | 罕见的 |
| 典型全色盲 | 非常罕见的 | 非常罕见的 |
在Cole (1972), Pease(1998)和Wright(1952)的估计之后修改*
根据受影响的锥体光色素,可分为二色和异常三色。三个术语也被用来描述CVD是protan, deutan和tritan(来自希腊语protos Ð第一,deuteros Ð第二,tritos Ð第三;颜色视觉缺陷的顺序描述)。质子有较长波长锥体色素缺失或异常,多肽有中波长锥体色素缺失或异常,三萜类有较短波长锥体色素缺失或异常。(图17)。

deutan(红-绿)CVD的遗传模式为性别连锁隐性遗传,而tritan CVD为常染色体显性遗传。在临床中,CVD的遗传学尤为重要,因为患者常常渴望了解他们为什么是CVD,以及他们是否会把CVD传给他们的孩子。一段时间以来,人们认为先天性遗传性三视并不存在,因为报告的病例太少。此外,三叉戟样色觉缺陷与疾病有关,因此必须区分获得性和先天性三叉戟样色觉缺陷。先天性遗传性三视的存在最初是由家庭研究确定的(例如,Henry et al., 1964;科尔等人,1966;Smith et al 1973),随后被分子遗传学证实(nathan et al., 1986a,b)。
使用CIE图表开发诊断色觉测试。
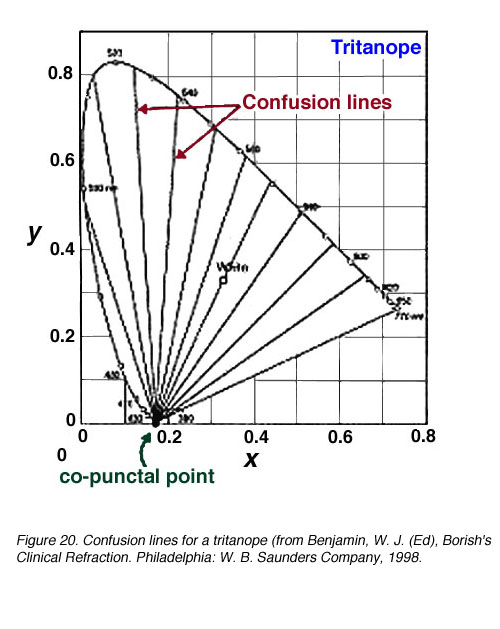
由于缺少一种光颜料,重色仪的辨色能力受到限制。因此,当谈到颜色搭配时,某些颜色会与另一种颜色混淆。混淆线是色度图上的连接点,对于二色仪来说,它们的颜色是相同的。混淆线的数量也提供了波长变化量的信息(DL)在区分另一种颜色之前(当遇到下一条混淆线时)。所有的混淆线都汇聚到一个点上,这个点被称为同颅点(图18)。

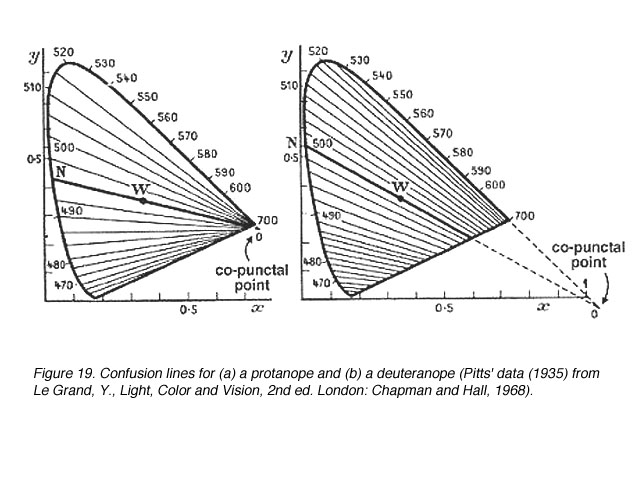
图19显示了一个丙烷烷和一个氘烷的混淆线。一个原嘌呤有17条混淆线,一个氘嘌呤有27条混淆线。因此,由于波长变化较小(DL步)可以辨别。图20显示了三角面的混淆线。
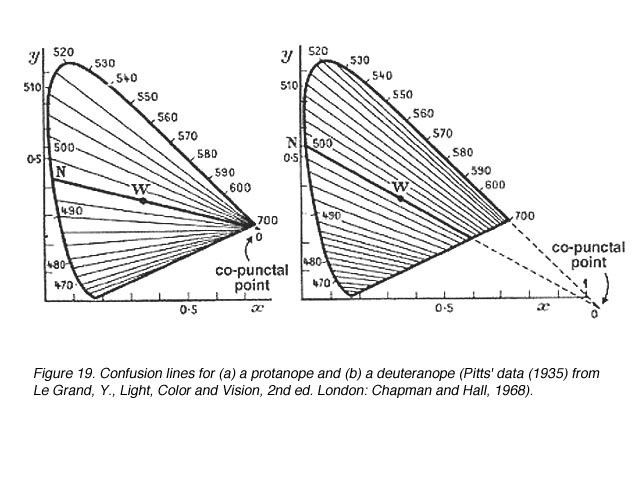 图19所示。(a)一个原面和(b)一个后面的混淆线(皮茨的数据(1935)来自Le Grand, Y.,光,颜色和视觉,2nd伦敦:查普曼和霍尔,1968年) |
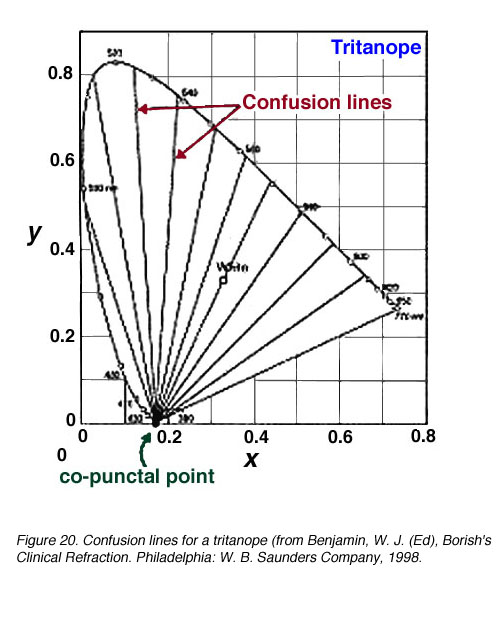 图20。三面书的混淆线(来自Benjamin, W. J. (Ed), Borish的临床折射。费城:W. B.桑德斯公司,1998 |
{kind=link}
{kind=link}
混淆线形成了许多色彩视觉测试的基础,如法恩斯沃斯D-15面板和石原伪同色板。假同色平板试验也常用于临床筛查色觉缺陷。颜色是根据混淆线精心选择的。临床上最常用的假等色板是石原等色板(用于筛查红绿色视力缺陷)和Tritan (F-2)板。
假同色印版有四种设计方法:
- 变换版:正常色觉的人看到一个图形,CVD的人看到另一个图形(图21a)。
- 消失版:色觉正常的人看到的图像,而CVD的人看不到(图21b)。
- 隐数字板:具有正常色觉的人看不到数字,而CVD则能看到数字(图22a)。
- 诊断板:设计用于有彩色视觉缺陷的正常受试者看到一个数字比另一个数字更容易(图22b)。
 图21。(a)石原的变形板。正常人应该看3个,而心血管疾病患者应该看5个。(b)石原消失的板块。正常应该看到73,而CVD将不能正确读取数字 |
 图22。(a)石原藏指盘。正常人应该看不到任何东西,而心血管疾病患者应该看到5个。(b)石原号诊断板。正常人应该能看到2和6。多多安型色觉缺陷者应该更容易看到2个,而多多安型色觉缺陷者应该更容易看到6个 |
{kind=link}
{kind=link}
石原色觉测试是一种筛选测试,石原色觉的失败标准通常是四个或更多的盘子。进一步的色觉测试将需要有信心地诊断色觉缺陷的类型。另一个有用的筛板试验是法恩斯沃思F-2筛板(皮斯,1998年)。有许多颜色视觉测试可用于筛查和诊断,其中只有少数将在这里讨论。D-15面板的颜色也从CIE图表中仔细选择,所以它们都是等发光的(也就是说,具有相同的值),如图23所示。
 图23。法恩斯沃斯面板D-15的颜色色度坐标(来自Benjamin, W. J. (Ed), Borish的临床折射。费城:W. B.桑德斯公司,1998)
图23。法恩斯沃斯面板D-15的颜色色度坐标(来自Benjamin, W. J. (Ed), Borish的临床折射。费城:W. B.桑德斯公司,1998)
患者被要求根据与试点颜色帽的相似度按顺序排列15个颜色帽(图24)。

颜色视觉缺陷的类型可以从他们的帽子的排列来检测。这些颜色帽以一种特殊的方式排列,这是由于位于混淆线上的颜色的混淆(图25)。在Panel D-15测试中失败的标准是两个或两个以上的主要交叉(即大于两个帽的误差)。多言、多言、三言会根据其混淆线产生特征误差(交叉)。杆状单色体为色盲,其Vl峰约为507 nm。他们根据D-15帽的scotopic反射率来排列D-15帽。

图25。D-15型法恩斯沃斯面板是由有各种颜色视觉缺陷的患者产生的。杆单色结果被理想化以说明沿5-14的scotopic轴。作为一种规则,杆单色仪给出的结果变化不定,交叉误差沿5-14轴下降的趋势
其他排列色彩测试包括L 'Anthony 's去饱和面板D-15,饱和H-16和法恩斯沃斯100色相测试。低饱和度的D-15在获得性疾病和轻度先天性缺陷的早期诊断中特别有用。试验管理的顺序包括用伪等色板试验(如石原板和F-2板)筛查色差。如果这两项测试中有一项失败,就意味着要进行D-15面板测试。没有通过D-15检查的患者被称为中度到重度异常重色谱仪,或重色谱仪。H-16检查失败的患者是重色谱仪或非常严重的异色色谱仪,而中度异色色谱仪将通过H-16。经过D-15区的受试者,可以进入饱和的D-15区,可以检测到轻微的毛锥体异常。非常轻微的异常三色视可能失败,也可能不会失败,可以用奈格尔异常镜进行诊断。
Farnsworth 100-Hue是另一个排列测试(图26)。不像上面提到的测试,颜色是专门选择靠近混淆线,Farnsworth100-Hue是一个辨别测试。

图27显示了100色相的颜色在色度图上的位置。颜色的选择具有相同的蒙塞尔值和色度。最初有100种色调,但法恩斯沃斯删除了15种,以使系列更加统一。Farnsworth 100-Hue的性能是通过计算总误差评分来评定的。

图27所示。色度图上的100色相的颜色。C点代表使用标准光源C的测试条件,W点是等能量白(来自哈特W. M. Jr,获得性色差。Surv角膜切削1987;32: 10)
灯测试自19世纪以来一直被用作评估色彩视觉的手段,尤其是职业原因。灯笼测试模拟彩色信号灯。他们通常呈现成对的红、白、绿灯,因为这些是在海上和空中航行中使用的信号颜色,并且要求受试者说出这些颜色的名称。有许多不同的灯笼测试,它们呈现的困难程度差别很大。困难的程度取决于刺激的规模和强度(参见Cole和Vingrys 1982年的回顾)。目前使用的灯笼是Farnsworth灯笼,现在被Optec 900、Holmes Wright A型和B型灯笼以及Beyne灯笼所取代(图28)。
灯笼测试通常被二色视者和异常三色视者失败,他们的缺陷严重到足以导致他们无法通过法恩斯沃思D15测试,但在轻度异常三色视者中,识别小彩色信号光的能力相差很大。通过D15测试或在异常镜上的小范围并不一定意味着信号光的颜色可以被识别(Cole和Maddocks, 1998)。

参考文献
Baylor DA, Nunn BS, Schnapf JL。猕猴竹杆的光电流、噪声和光谱灵敏度。杂志。1984;357:575 - 607。PubMed] [免费全文在PMC]
本杰明WJ,编辑器。Borish临床折射。费城:W. B.桑德斯公司;1998.
Bowmaker JK, Dartnall HJ。人眼视网膜视杆和视锥色素。杂志。1980;298:501 - 511。[PubMed] [免费全文在PMC]
三视症的表型变异。视觉研究》1966;6:301 - 313。
彩色视力异常的障碍。大屠杀J Optom。1972;55:304-310。
科尔BL,温格雷斯AJ。彩色视觉灯笼试验的调查与评价。Am J Optom Physiol Opt. 1982; 59:46 - 374。[PubMed]
科尔BL,温格雷斯AJ。谁没通过灯笼测试?医生角膜切削。1983;55:157 - 173。[PubMed]
科尔·BL,麦多克斯·JD。临床色彩视觉测试可以用来预测法恩斯沃斯灯笼测试的结果吗?视觉研究》1998;38:3483 - 3485。[PubMed]
眼睛。卷2。伦敦:文献出版社,1962
恒河猴视网膜神经节细胞的功能特性。杂志。1975;251:167 - 95。[PubMed] [免费全文在PMC]
德瓦卢瓦,阿布拉莫夫一世,雅各布斯GH。LGN细胞反应模式的分析。J Opt Soc Am. 1966; 56:966-977。[PubMed]
Deeb, SS(2004)色觉缺陷的分子遗传学。神经科学21,191-196。[PubMed]
猴神经节细胞锥细胞形成机制的鉴定。杂志。1968;199:533 - 547。[PubMed] [免费全文在PMC]
颜色:数据和理论(第15章)和依赖于波长的鉴别(第12章)。作者:格雷厄姆·CH,编辑。视觉和视觉知觉。纽约:约翰·威利父子公司;1965.
Grassmann HC。Zur theory orie der farbenmischung。安phy。1853;89:69 - 84。
哈特WM。获得了色觉障碍。Surv角膜切削。1987;32:10-31。[PubMed]
Henry GH, Cole BL, Nathan J.先天性三视的遗传与广泛的谱系报告。安·胡姆·吉内1964;27:19 - 231。[PubMed]
郝林KEK。Zur Lehre vom Lichtsinne。这是一个伟大的国家。数学-自然科学schaftliche班,Wien, 3。1872; 66:5-24。
色彩视觉的对立过程理论。Psychol启64:384 1957;404年。[PubMed]
Le Grand Y.浅色和视觉。第二版。伦敦:查普曼和霍尔1968年。
马克斯WB,多贝尔WH,麦克尼科尔JR.单一灵长类视锥细胞的视觉色素。科学。1964;143:1181 - 1183。[PubMed]
万豪FHC(1962)色彩视觉:色彩匹配。第13章。在:戴维森,H. (ed),眼睛,第二卷.伦敦,学术出版社。
摩西RA,哈特WM。阿德勒的眼睛生理学。临床应用。第八版。圣路易斯:C. V.莫斯比公司。1987
nathan J, Piantanida TP, Eddy RL, Shows TB, Hogness DS。人类色觉遗传变异的分子遗传学。科学。1986;232:203 - 210。PubMed]
内森斯J,托马斯D,霍格内斯DS。人类色觉的分子遗传学:编码蓝、绿、红色素的基因。科学。1986;232:193 - 202。PubMed]
Patterson EJ, Wilk M, Langlo CS, Kasilian M, Ring M, Hufnagel RB, Dubis AM, Tee JJ, Kalitzeos A, Gardner JC, Ahmed ZM, Sisk RA, Larsen M, Sjoberg S, Connor TB, Dubra A, Neitz J, Hardcastle AJ, Neitz M, Michaelides M, Carroll J. (2016) x -连锁视锥功能障碍和红绿色视力缺陷患者的视锥光感受器结构。《眼科科学》57(8):3853-63。[PubMed]
皮斯PL。(1998)。色彩视觉(第9章)。在:本杰明WJ,编辑。Borish临床折射。费城:W. B.桑德斯公司。
拉什顿哇。protanope的一种锥形色素杂志。1963;168:345 - 359。[PubMed]
拉什顿哇。正常人和色觉缺陷者视杆和视锥色素密度测定。角膜切削投资。1966;5:233 - 241。[PubMed]
Smith DP, Cole BL, Isaacs A.无神经视网膜疾病的先天性三视。Ophthal投资。1973;12:608 - 617。[PubMed]
中心凹锥光色素在400 ~ 500 nm范围内的光谱灵敏度。视觉研究》1975;15:161 - 171。[PubMed]
Svaetichin G.单锥光谱响应曲线。物理学报。1956;39:17-46。[PubMed]
冯·赫姆霍尔兹。Handbuch der physiology ischen Optik。[English translation in MacAdam DL.(1970). Sources of color science. Cambridge (MA): MIT Press; 1970] Hamburg: L. Voss. 1866
赖特WD。三视症的特点。J Opt Soc Am 1952; 42:509-520。[PubMed]
关于光和颜色的理论。Philos Trans R Soc Lond. 1802; 92:12-48。
灵长类动物视网膜神经节细胞的蓝色敏感锥机制特征。视觉研究》1981;21:1605 - 1609。[PubMed]
网站ghr.nlm.nih.gov遗传学家参考《色觉缺陷》。国家医学图书馆。
最近更新:2007年6月5日。
作者

迈克尔Kalloniatis1958年出生于希腊雅典。他获得了墨尔本大学的视光学士学位和硕士学位。他在休斯顿大学视光学院获得博士学位,以研究猴子视觉系统中的颜色视觉处理。beplay体育公司博士后培训在休斯顿的德克萨斯大学继续,师从罗伯特·马克博士。正是在这一时期,他对视网膜神经化学产生了浓厚的兴趣,但他也保持着一个活跃的视觉心理物理学研究实验室,专注于色彩视觉和视觉适应。在他最近搬到新西兰之前,他曾是墨尔本大学视光与视觉科学系的教员。Kalloniatis博士现在是奥克兰大学视光与视觉科学系的Robert G. Leitl视光教授。电子邮件:m.kalloniatis@unsw.edu.au
作者
查尔斯灾区1974年出生于越南参苴。他在墨尔本接受教育,并于1996年获得墨尔本大学的视光学位,随后在维多利亚视光学院进行临床住院治疗。在此期间,他完成了研究生培训,并获得了临床验光研究生文凭。他的专业领域包括低视力和隐形眼镜。在他担任验光师期间,他与Michael Kalloniatis博士合作,教授验光学生,并组装了“独眼”。Cyclopean Eye是一个基于网络的交互式单元,用于验光学本科生的视觉科学教学。他目前是墨尔本大学视光与视觉科学系的私人验光诊所和访问临床医生。