1简介
“生物学中没有任何东西是有意义的,除非从进化的角度来看”(1)。
狄奥多西·多布赞斯基(Theodosius Dobzhansky)的洞察力尤其适用于试图理解我们视杆和视锥感光器的本质,以及视网膜的组织。除非我们了解这些细胞和结构是如何通过数亿年的进化产生的,否则我们就很难理解它们的形态和分子结构,或者能够回答“视网膜为什么会这样?”这个反复出现的难题。除了为我们视网膜的排列提供理论基础之外,对我们眼睛及其视锥细胞和视杆细胞的进化的研究也非常令人满意,它为诸如“我们的眼睛是何时、如何起源的”等问题提供了潜在答案。以及“为什么我们和其他脊椎动物的眼睛和(比如)昆虫的眼睛如此不同?”
在脊椎动物眼睛进化的过程中,明显缺乏被保存下来的过渡形态,这为“神创论者”提供了常年的素材。但是,正如查尔斯·达尔文所解释的,
“如果从一只完美而复杂的眼睛到一只非常不完美而简单的眼睛有无数个等级,每一个等级对它的拥有者都是有用的,可以证明存在……如果器官的任何变异或修饰在生活条件不断变化的情况下对动物有用,那么,相信一只完美而复杂的眼睛可以由自然选择形成的困难,尽管我们的想象力无法克服,就很难被认为是真实的。”
这篇文章的目的之一是记录在脊索动物和脊椎动物进化过程中,光感受器、光转导级联和视网膜中的这种分级的证据。第二个主要目标是为一长串事件构建一套“场景”;在这方面,“情景”一词在字典中的意思是“事件的假定顺序或发展”。
历史上,研究眼睛进化的三个主要途径被利用:检查化石记录中的眼睛,检查现存物种的眼睛结构,以及检查胚胎发育。最近,利用分子证据开发出了一些强有力的新途径;例如,跨现存物种的比较分子遗传学,以及进化和发育分析的结合(进化-进化-进化方法)。这篇文章集中在现存的脊索动物的眼睛上,并从形态学、胚胎学和分子特征中研究眼睛进化的线索。在这样做的过程中,它建立在Lamb等人(3,4)提出的场景之上。
关于光转导和光接收进化的其他方面的最新综述,参见Arendt等人(5),Vopalensky & Kosmik (6), Larhammar等人(7),Shichida & Matsuyama (8), Kusakabe等人(9),Collin等人(10),Fain等人(11),Porter等人(12)和Nilsson(13)。关于整个动物王国(而不是主要在脊索动物中)进化出的眼睛类型的描述,请参阅Schwab的插图丰富的书(14)。关于脊椎动物感觉系统和大脑的进化,见Butler(15)和Butler & Hodos(16)。
经常被问到的一个问题是“眼睛和光感受器独立进化了多少次?””这个问题的答案因人而异,这取决于一个人对“独立”的理解。下面我们将看到,刺胞动物、原口动物和后口动物的共同祖先已经具备了构成眼球和/或视网膜所需的绝大多数成分;例如,它已经拥有转录因子、生长因子、视蛋白、光感受器细胞(纤毛型和微绒毛型)、色素细胞和神经元等。使用这组通用的工具,不同的事件发生在不同的谱系中,导致了非常不同的视角。在某些原口动物中,一个单眼多次复制形成复眼;在我们人类的谱系中,形成了一个广泛的光敏视网膜,并在其前面发展了一个单一的光学元件。
虽然由此产生的眼睛彼此之间有根本的不同,但事实证明,它们所基于的光感受器彼此之间非常相似,而且确实源自一个共同的祖先类型,因此我们现在可以断然否定Salvini-Plawen & Mayr(17)的主张,即“光感受器至少起源于40个不同的系系,但可能多达65个或更多”。
正如Jacob(18)非常恰当地指出的那样,重要的是要认识到,进化是通过“修补”现有的东西来进行的,而没有任何总体的“目的”(比如产生视觉)。在光转导、光感受器和视网膜的进化过程中,大量此类修补的例子将在前面的章节中变得明显;参见Goldsmith(19)。
2起源
作为背景,以下各部分依次简要概括:脊椎动物的起源、脊椎动物眼的起源、视蛋白的起源和感光细胞的起源。
脊椎动物的起源
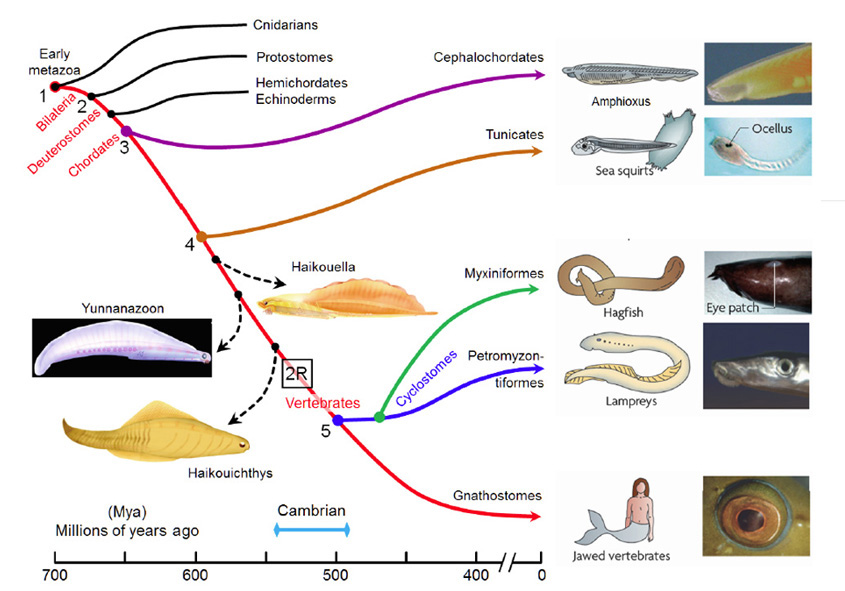
图1概述了脊椎动物的起源,时间跨度从700至400万年(百万年前);这张图主要说明了与脊椎动物进化相关的现存类群,虽然也显示了三个感兴趣的灭绝类群(使用虚线)。尽管人们对分支发生的顺序有广泛的共识,但对各种分支发生的时间却不太确定。分子证据表明,许多分支发生的时间比化石记录中记录的要早得多,图1所示的时间是根据Erwin等人(20)报道的分子和化石证据最近的调和得出的。在接下来的讨论中,我们将从我们的直系祖先的角度来描述这些分支,如红色曲线所示,将各种重要的分支编号为#1到#5。
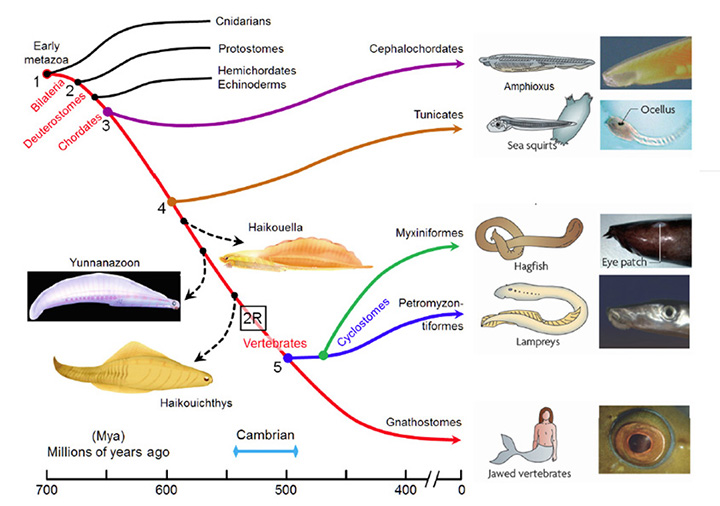
大约在700米左右(#1),早期的真目动物是我们的远祖,它们分为刺胞动物(如水母、珊瑚等)和我们自己的双侧对称动物(Bilateria)。然后,在665年左右(#2),绝大多数现存无脊椎动物物种(原口动物)的祖先从我们的后口动物谱系中分离出来。几百万年后,半足类动物和棘皮类动物(如海胆)的共同祖先从我们的祖先中分离出来。不久之后,大约在655玛雅(#3),头脊索类动物(如文昌鱼)分裂了——从那次分裂开始(最晚),我们的祖先已经是脊索类动物了。大约又过了5000万年,大约在600米(#4),被囊动物(例如海鞘)分化了,然后又过了1亿年,才发生了下一次分裂,后代得以存活下来,当时现存的无颚脊椎动物(agnathan)的祖先从我们自己的有颚脊椎动物谱系(颌口动物)中分化出来,大约在500米(#5)左右。从这种分化开始(可能在此之前),我们的祖先就已经是脊椎动物了。
之前的1亿年间,也就是4 - 5年之间(600-500米),不仅对脊椎动物眼睛的进化很重要,而且更普遍地说,它是身体结构进化中异常创新的时期。然而,令人沮丧的是,有两个因素使对那段时期发生的转型的分析复杂化。首先,在那一亿年的时间跨度中,从我们自己的谱系中分离出来的众多中间形态中,没有一个存活到今天。图1显示了一些灭绝物种的重要例子,但现存物种之间存在着巨大的差距。其次,软组织在化石记录中保存得很差,因此很少能从已知的灭绝物种中获得脊索动物眼睛过渡形态的线索。
尽管有这些困难,关于脊椎动物的眼睛进化,我们可以推测出相当多的东西,(我)来自对现存动物眼睛的比较分析,(2)来自胚胎眼发育的分析,以及(3)来自光感受器和视网膜的分子遗传分析。但是,在我们详细研究现存脊椎动物的眼睛之前,如果我们简单地考虑一下脊椎动物眼睛的起源、视蛋白光色素的起源和感光细胞的起源,将有助于建立一个场景。
脊椎动物眼的起源
来帮助回答这个问题“脊椎动物的眼睛是什么时候出现的?”人们可以对现存的脊索动物的眼睛进行有益的检查——下颌脊椎动物、圆口动物、被囊动物和头脊索动物。
喋喋不休的脊椎动物。所有现存的下颌脊椎动物(如鱼类、四足动物、鸟类)的眼睛都非常相似,而且似乎是按照一个共同的平面构造的。因此,我们可以肯定,生活在420万年左右的所有现存颌骨脊椎动物的最后一个共同祖先,拥有“脊椎动物式的眼睛”。但我们能更深入地探究过去吗?
七鳃鳗。七鳃鳗的祖先在500米左右与我们的祖先发生了分化(图1),然而七鳃鳗的照相机式眼睛与颌鱼和其他颌脊椎动物的眼睛极其相似。因此,七鳃鳗的眼睛有一个晶状体、一个虹膜和一组六块眼外肌肉,这些肌肉与颌类脊椎动物的眼外肌肉非常相似(第3节)。此外,七鳃鳗的视网膜结构与脊椎动物的视网膜结构非常相似,五类同源神经元(光感受器、水平细胞、双极细胞、无分泌细胞和神经节细胞)分布在三个主要核层和两个丛状层。南半球七鳃鳗(lamprey Geotria australis)具有五种形态的视网膜感光细胞和五种视蛋白,每一种视蛋白都与下颌脊椎动物的视网膜视蛋白密切相关(详见第6节);然而,北半球的七鳃鳗物种已经失去了几个视蛋白类。
鉴于七鳃鳗和下颌脊椎动物的眼睛极为相似,几乎可以肯定的是,这些类群的最后一个共同祖先拥有照相机型的眼睛,与现存的七鳃鳗和颌口动物的眼睛大致相当,因此,大约500年前脊椎动物的眼睛就已经存在了。然而,我们不能完全否认这样一种可能性:即使七鳃鳗和下颌脊椎动物的最后共同祖先拥有必要的基因,它的眼睛可能表现出一种比完全发育的照相机式眼睛更原始的形式,而且七鳃鳗和颌口动物都通过某种程度的趋同进化完善了眼睛的物理表现形式。
八目鳗类鱼.盲鳗的“眼睛”是一种特殊情况,我们将在第三节详细讨论。尽管盲鳗是七鳃鳗类祖先的后代,但它们的“眼睛”比七鳃鳗的眼睛表现出更基本的形式,而且人们有充分的理由认为,这些特征是从眼睛进化的早期阶段保留下来的。
被囊动物和无头类。在脊索动物类群中,在长尾类动物(七鳃鳗和盲鳗)之前就已经分化了,没有一种存活到今天的脊索动物拥有可以被恰当地描述为眼睛的器官。头脊索类文昌鱼有好几组感光细胞,幼虫有一个单眼。在这两种情况下,有c视蛋白的纤毛光感受器都存在,并且看起来与我们自身的视锥和视杆细胞同源。虽然它们的光感受器和视蛋白为脊椎动物光感受器的进化提供了重要线索,但这些原始生物显然在脊椎动物相机眼进化之前就已经分化了。
一些令人感兴趣的灭绝脊索动物。三种已经灭绝的早期脊索动物在图1中有图示(图表来自Chen(21))。这是已知的最早的脊索动物,它们拥有的结构在现存脊椎动物中是由迁徙的神经嵴组织形成的,因此它们被Chen和同事命名为“嵴动物”或Cristozoa。这些前颅顶化石仅在云南下寒武统地层中发现过;它们大约3厘米长。在这三种物种中,Haikouella生长状况有最基本的形式,和Yunnanazoon lividum看起来更发达,而Haikouichthys ercaicunensis是最发达的,似乎是头盖骨的过渡形式。据报道,每一个化石物种都有成对的眼睛,直径分别约为0.3、0.6和0.4毫米,但不幸的是,这些软组织的细节保存得太少,以至于无法描述眼睛的内部特征——甚至,例如,它们是否有晶晶体。
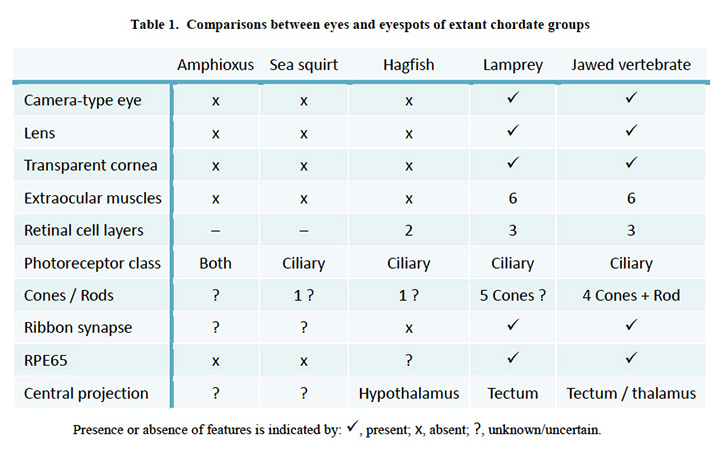
脊索动物眼特征的比较。表1比较了上述现存脊索动物的眼睛(或眼斑)的一些特征。毫无疑问,我们与七鳃鳗的最后一个共同祖先(约500米,图1、#5)拥有一个完全成熟的“脊椎动物式的相机眼”,而再往前追溯,我们与海鞘的最后一个共同祖先(约600米,图1、#4)除了一个简单的眼点之外,可能还有更复杂的东西,这似乎是不可思议的。
脊索动物和其他门的眼睛进化的关键时期。很明显,第4到第5段,600-500米之间,对脊椎动物眼的进化至关重要。此外,身体外观最深刻的变化发生在那个时期的末期,也就是寒武纪身体形态大爆发的时期,这似乎是合理的。大约在同一时间,眼睛也在许多其他门中进化,通常具有截然不同的物理形式(14,22)。
最近发现的保存非常完好的早寒武纪(约515米亚)眼睛化石表明,一些最早的节肢动物已经拥有含有数千个小眼的复眼。Paterson等人(23)报道Anomalocaris眼睛直径至少12毫米,包含超过16000个小眼。这些动物的身体可达90厘米长,被认为是自由游动的顶端掠食者,现在已被证实拥有具有高空间分辨率(~1°)潜力的复眼。我们自己的祖先眼睛发育迟缓,他们可能被视觉引导的原口动物捕食了数千万年。
眼睛进化的动力是什么?在分析寒武纪感觉系统的出现时,Plotnick等人(24)提出了两个使获得空间视觉变得非常有价值的因素,一是寒武纪景观中空间复杂性的急剧增加,二是自由游动的生物需要导航。来自空间视觉的光流信息至少为后一种需求提供了部分解决方案。在这些压力之上,当然还有发现猎物和躲避捕食者的需要。
视蛋白的起源
视蛋白及其主要的分类(25)在后生动物进化的早期就出现了。在本文中,“视蛋白”一词仅指“2型动物视蛋白”,而不是细菌的“1型微生物视蛋白”或藻类的“通道视蛋白”,这两种视蛋白都是不相关的,似乎是由趋同进化产生的。睫状体视蛋白的系统发育将在第5节和第6节讨论脊索动物和脊椎动物视网膜的系统发育,但目前的问题是:祖先视蛋白是如何起源的?而且它多元化的最初阶段是什么?.为了解决这些问题,通过分析自2007年以来可用的一些刺胞视蛋白序列(12,26 -31)获得了重要的线索。
动物视蛋白从g蛋白偶联受体(GPCRs)的“GRAFS”超家族的同名“视紫红质家族”中进化而来,已知该超家族起源于一种古老的真核生物,存在于真菌分化之前(32)。最近,封建等人(30)分析了视蛋白的系统发育,并提出了视蛋白的早期起源方案。他们发现,视蛋白的近亲存在于包括脊椎动物褪黑激素受体的谱系中。然而,对于无脊椎动物中相应的GPCRs,配体还没有被识别出来,因此尚不清楚视蛋白谱系分化时的祖先配体可能是什么。
封建等人(30)的分析的一个潜在问题是,它依赖于(未经证实的)刺胞动物中r -视蛋白的存在,但这个问题似乎已经被一项独立的、几乎同时进行的对珊瑚视蛋白的研究(31)解决了,该研究清楚地确定了r -视蛋白的存在。以下关于动物视蛋白早期起源的情景,如图2B所示,是建立在Feuda等人(30)的报告基础上的,在这里作为导致光感受器产生的一系列情景/假设的第一个:
a- 1)第一个视蛋白的先驱是在一个古老的后生动物中通过GPCR的复制而产生的,在类似阿米巴的扁平动物分化之前。
A-2)该前体蛋白在第7跨膜螺旋中不具有视网膜结合赖氨酸(' K296 ') (30);这表明,与传统GPCRs中的配体一样,视黄醛配体通过非共价结合的方式占据了内腔,在图2B中,这种前视蛋白被称为“视黄醛受体”。柔虫Trichoplax有视蛋白的同源物(封建等人,2012年命名为视蛋白),同样没有视网膜结合赖氨酸残留。
A-3)在受体的第7跨膜段内获得适当位置的赖氨酸残基,使视黄醛配体以共价结合。最初,希夫碱键很可能没有被酸化,所以分子会被紫外线吸收。获得适当位置的负电荷残基(如E181)允许键被质子化,从而产生原始视蛋白,并使吸收峰转移到“可见”光谱。
A-4)对于大多数视蛋白(脊椎动物视蛋白除外),这种视蛋白激活的后视视紫红质状态是热稳定的,可以通过光逆转到视紫红质状态。因此这种蛋白质可能不需要11-的来源独联体视网膜和可替代利用所有-反式视网膜很好。
A-5)随后,在扁平动物分化和刺胞动物从两侧动物分化的相对较短的时间内,出现了两个最早视蛋白的复制。因此,图2B中显示的所有重复都发生在图1中编号的第一个分支之前(即在#1之前)。
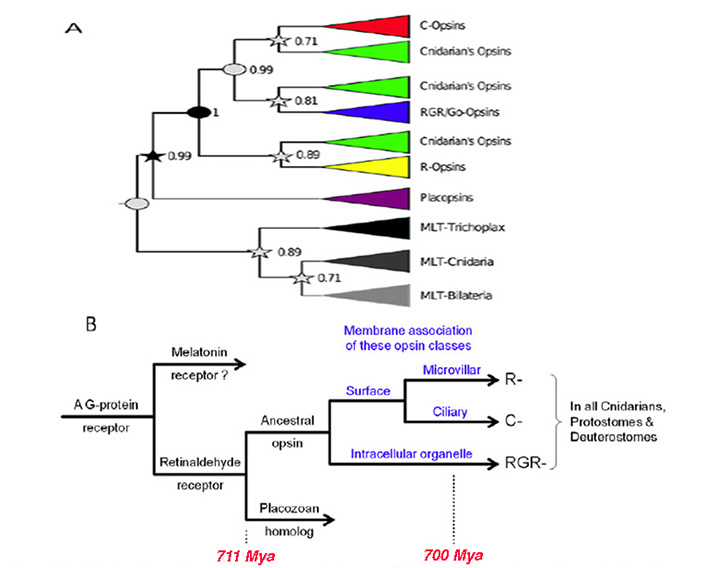
假设视蛋白类型和膜类型之间的联系.视蛋白类和光感受器类共同进化的一个因素可能是不同视蛋白与膜的不同区域的优先结合,如图2B所示。因此,视蛋白早期进化的假设情景扩展如下:
A-6)在第一次复制事件后出现的两种视蛋白变体可能优先转移到亚细胞细胞器的膜和表面膜。这些变异将分别导致现代视蛋白的RGR-分裂和C-/R-分裂。
A-7)在C-视蛋白和r -视蛋白之间产生区别的复制事件发生后,这两种变体分别进入纤毛膜和微绒毛膜。在图2B中,这种复制显示在上一点提到的复制之后发生,但目前还不能可靠地区分这对复制事件发生的顺序。
a -8)随后,表达C-和r -视蛋白类的细胞通过一个称为“分工”的过程变得彼此不同(5,33),导致(a)拥有C-视蛋白的纤毛光受体和(b)拥有r -视蛋白的微绒毛光受体;见下一节。视蛋白的第三种变体,rgr -视蛋白,倾向于在细胞内细胞器的膜中表达,可能作为前两类光感受器中的额外视蛋白。
在进化的后期,进一步的分工发生了,因此(例如)rgr -视蛋白可以在单独的细胞中表达。这就解释了为什么一方面,乌贼的光感受器在它们的微绒毛膜中含有r -视蛋白,同时在它们的细胞内细胞器中含有视色素(rgr -视蛋白);另一方面,脊椎动物的视锥和视杆细胞在它们的外节段中只含有c -视蛋白,而RPE细胞在内质网中只含有rgr -视蛋白。
光感受器细胞类型的分类与多样性
一种广泛流传的观点是,光感受器细胞分为两种:无脊椎动物(原口动物)眼睛中的横纹肌光感受器和脊索动物眼睛中的纤毛光感受器。然而,光感受器的类型与门的刚性联系,以及光感受器的刚性分为两类,都不能被支持。
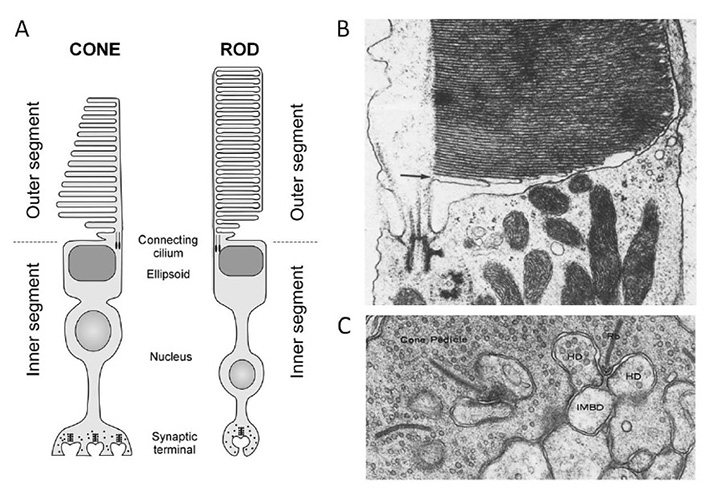
睫状肌和横纹肌的分类。根据含有视蛋白的区域形成广阔表面积的方式,埃金和他的合作者辨别出两类光感受器毛状的形态,以脊椎动物的杆状和锥状感光器为例,其中膜扩张形成修饰的纤毛,从经典的9+0非运动轴突结构辐射出来rhabdomeric形态,以昆虫的光感受器为例,膜的扩张是微绒毛以高度有序的方式排列的(33);参见下面的图3和图4。
在很大程度上,基于Eakin的分类,提出了三种理论来解释光感受器类型的进化。Eakin自己提出了一个后来被称为光感受器进化的双系模型,其中光感受器横纹肌系出现在原气孔中,作为纤毛光感受器祖先系的变体(34)。Eakin承认他的模型有一些例外(比如带有微绒毛光感受器的后口动物),但他提出这些例外是通过独立进化产生的。
在Eakin模型的一种变体中,Vanfleteren & Coomans(35)解释横纹肌光感受器代表了祖先纤毛光感受器的一种改良形式,其中纤毛结构诱导微绒毛膜的形成,而纤毛结构往往随后退化;其他作者称之为光感受器进化的单系模型。与上述两种模型相反,Salvini-Plawen & Mayr(17)提出光感受器至少在40种情况下独立进化,他们称之为光感受器的多系起源。事后看来,埃金和范弗雷特伦&库曼斯的模型都有可取之处。
光感受器类型的多样性。在动物王国中,光感受器类型的多样性确实是惊人的,以至于可能在大多数生物体中,光感受器的形态不能完全归为纤毛和横纹肌两类。对于发生在不同生物体中的光敏结构的范围的概述,感兴趣的读者可以参考Vanfleteren(36)提供的光感受器形态学的综合摘要。除了传统的“纤毛”和“横纹肌”结构外,还会发生许多种类的膜细化。
此外,如图3(36)所示,纤毛膜和微绒毛膜经常同时出现在单一类型的光感受器中。例如,在发育过程中,微绒毛光感受器通常表现出中心粒等纤毛结构(图3A),尽管在原口动物的大多数情况下,这些纤毛是短暂的。
重要的是,在后口动物(甚至是脊椎动物)的光感受器中也能看到这种共生现象。文昌鱼的微绒毛光感受器中存在纤毛;因此,黑塞细胞有一个9+0纤毛,而约瑟夫细胞有两个纤毛(37)。脊椎动物的纤毛光感受器通常具有微绒毛,这些绒毛来自于纤毛内段的远端区域,并与纤毛外段密切相关(图3B3);然而,这些微绒毛不表现出横纹肌光感受器的典型组织,与入射光成直角,而是纵向排列的。后口动物中的第三个例子是在半足形虫中发现的,橡子虫幼虫的脑眼含有感光细胞,既拥有发育良好的纤毛,又拥有大量紧密排列且与轴成直角的微绒毛(38)。
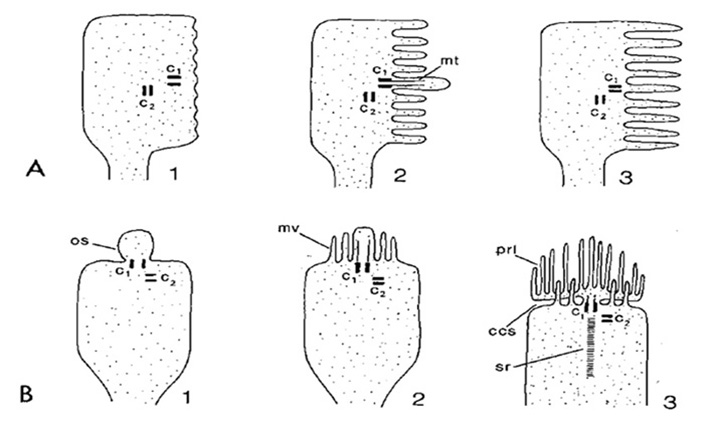
最后一个具有启发性的例子来自海洋腹足动物,Aporrhais pespelecani.在幼虫的眼睛中,光感受器是纤毛的,但在变形时,纤毛光感受器又发育出微绒毛,从而在成虫的眼睛中转变为纤毛+微绒毛的混合光感受器(39);不幸的是,视蛋白的性质和转导级联在这些光感受器尚不清楚。
即使是极端的情况也是同源的。由于光感受器形态的多样性,我们可以把苍蝇的光感受器和脊椎动物的杆细胞的经典案例看作是光感受器结构类型的巨大等级的极端。但是,尽管这两种细胞类型之间的形态差异很大,但它们之间的同源性(图4)是相当显著的,正如Ready & Tepass(40)所指出的。两种细胞均由单一柱状上皮发育而来。两者都保留了一个附着带(ZA)区域,该区域将基底膜和根尖膜分隔开来,并连接相邻的细胞;在脊椎动物的视网膜中,这些细胞间的接触形成了外限制膜。在发育两种类型的光感受器时,在ZA顶端的膜上发育两个子结构域。第一个是一个丰富的支持域果蝇形成苍蝇柄,脊椎动物的光感受器形成内节。第二个更尖的子区域大量扩张形成光敏膜果蝇如微绒毛,在脊椎动物的光感受器中如纤毛囊或纤毛盘。
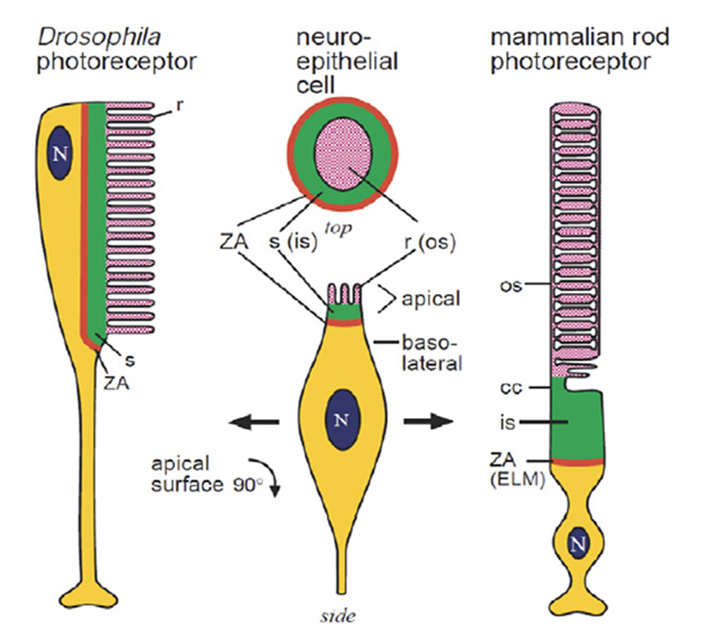
光感受器细胞起源的场景
从以上的概念中提炼出线索,我们提出了以下情景,即光感受器的两种主要形态变异的起源毛状的而且microvillar:
B-1)纤毛型代表光感受器的祖先类。这种祖先光感受器在纤毛膜上表达祖先视蛋白,在纤毛附近的胞体上也有微绒毛。
B-2)在后口动物和刺胞动物中,细胞膜纤毛区大量细化的机制进化。
B-3) r -视蛋白与c -视蛋白分离后(见上文),它们倾向于运输到微绒毛膜。
B-4)在双侧动物中,这些微绒毛膜的细化机制进化,导致了“纤毛”和“微绒毛”光感受器之间的主要区别。
B-5)在某些节肢动物和软体动物中,微绒毛的形成在高度组织性横纹肌的形成中达到了高潮,因此进化出了许多真正的“横纹肌”光感受器。
B-6)含有视蛋白的膜的许多不同的细化机制已经出现,这导致了许多不同种类的膜特殊化的进化,从而形成了不同生物体中大量的光感受器形态。
视蛋白类与转导级联的关系。视蛋白的种类与光感受器中g蛋白转导级联的性质密切相关。在脊椎动物中,c -视蛋白通常激活Gi家族的一个成员(包括Gt和Ggust),导致环核苷酸水平的调节和cngc(环核苷酸门控通道)的打开改变,从而产生电反应。相反,r -视蛋白激活Gq, Gq使用PLC(磷脂酶C)作为效应蛋白,TRP/C通道通常产生电活性。Plachetzki等人(27)提供的证据表明,祖先视蛋白很可能利用cngc来调节其电反应,r -视蛋白与TRP/C通道的连接很可能是随后出现的。视蛋白及其转导级联的共同进化的分子遗传学证据将在第8节中展开。
3.现存脊索动物眼睛中的睫状体光感受器
为了理解脊椎动物眼睛的进化,主要的挑战之一是解释在脊椎动物出现之前,古代脊索动物祖先的少数感光细胞和色素细胞可能进化成精致的视网膜的事件顺序。(与此密切相关的问题,在此不作讨论,但涉及必要的光学和运动器官以及适当的大脑区域的共同进化。)在考虑脊椎动物视网膜的进化时,我们现在将研究现存脊索动物的感光器官,从最简单的眼球开始,发展到脊椎动物的视网膜。对于每一个脊索类光敏器官的例子,我们将简要地检查该器官的一般特征,然后集中于它的纤毛光感受器。
在第7节中,我们将考虑为什么是纤毛光感受器而不是微绒毛光感受器在脊索动物和脊椎动物的眼睛中获胜。然后在后面的章节中,我们将研究哺乳动物视网膜的胚胎学发展,以及眼睛进化的分子线索。
头脊索动物的感光器官(文昌鱼)
Lacalli(41,42)描述了头脊索动物、文昌鱼的四个不同的光敏感区域(图5)。每个区域都沿着神经管,处于相当靠后的位置,在中线上(即不成对),没有任何类型的晶状体或其他成像设备。其中两个包含纤毛细胞(通常被认为是光感受器),而其他两个包含微绒毛光感受器。尽管后一种细胞过去常被称为“横纹肌”细胞,但它们实际上并没有表现出真正横纹肌的高度组织结构,最好将其称为“微villar”,如Gomez等人(43)及其同事所使用的那样。
纤毛细胞。“额眼”是一个微小的吻侧区域,包含一些纤毛细胞,被认为与脊椎动物的侧眼相似。“层状体”,在前眼后面不远,也包含睫状体细胞,在这种情况下,睫状体显示出非常广泛的层状膜;这个器官被认为是脊椎动物松果体的同源物。虽然层状体在幼虫中作为一个独特的器官存在,但在成虫中细胞似乎分散。到目前为止,这两种类型的纤毛细胞(前眼或板层体)都没有表现出对光的反应。
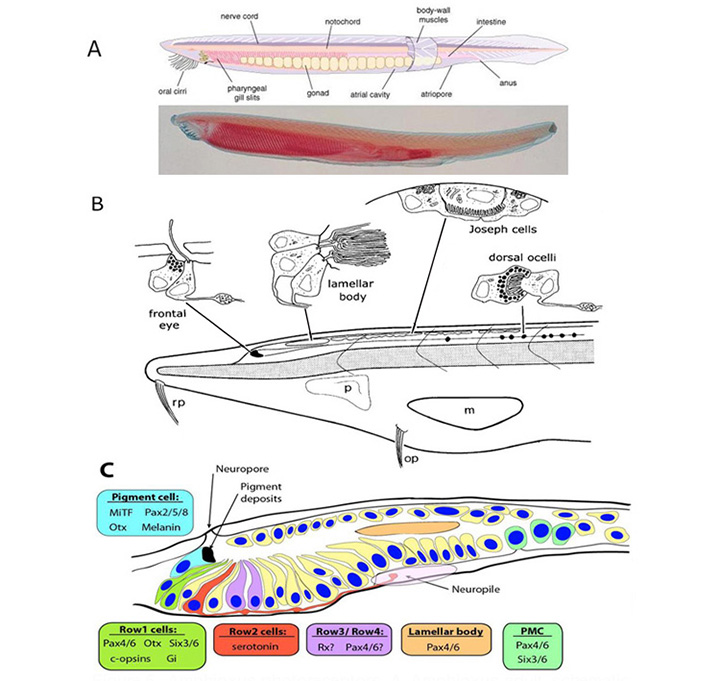
Microvillar感光细胞.再往尾端一点,发现了一组“约瑟夫细胞”,每一个都是一个微绒毛光感受器,再往尾端,发现了一链“背眼”或“黑塞器官”,每一个都包含一个部分被色素细胞包裹的微绒毛光感受器。最近,研究证实,这些细胞中的光转导与原口动物横纹肌光受体中的光转导非常相似,利用r -视蛋白黑视素(44)触发涉及Gq和PLC的经典“横纹肌”转导级联(44,46),这可能与脊椎动物视网膜内感光视网膜神经节细胞(ipRGCs)中的级联非常相似。
分子标记。最近,Vopalensky等人(47)在文昌鱼的前眼细胞中发现了许多表达的分子标记(图5C),他们得出的结论是,这一器官似乎确实与脊椎动物的侧眼同源。特别是前眼第一排的单睫状体细胞与锥体和杆状光感受器同源,与转录因子Pax4/6、Otx和Six3/6共同表达c -视蛋白;此外,抑制性gi型g蛋白α亚基的存在暗示了脊椎动物对光的OFF反应。第二排的细胞可能与视网膜神经节细胞同源;它们将轴突投射到神经泌,并且含有血清素,尽管到目前为止还没有足够的信息来确定它们可能与神经节细胞同源。最后,在黑色素含量、邻近感光细胞的位置和调控信号(Mitf、Otx和Pax2/5/8)方面,着色细胞似乎与脊椎动物RPE细胞同源。
先前提出的板层体与脊椎动物松果体器官的同源性是不被支持的,因为Otx和Rx都被证明是不存在的,同时也无法检测视蛋白。因此,尽管层状体的细胞明显表现出纤毛层状超微结构,但还没有证据表明它们是光感受器,而且该器官似乎与脊椎动物的眼睛或松果体不同源。
电生理学。文昌鱼的约瑟夫细胞和背单孔细胞最近已经被电生理技术研究,并被明确地确定为微绒毛光感受器(43),但额眼和板层体的睫状体细胞还没有被用这种方法研究过。
有被膜的单眼(幼虫Ciona而且Aplidium)
单眼。现存最接近脊椎动物的姐妹类群包括被囊动物,包括海鞘等Ciona intestinalis而且Aplidium constellatum.没有报道过无梗的成年海鞘具有任何类型的离散眼点,尽管分散的视蛋白表达细胞明显存在。然而,蝌蚪状的幼虫阶段(图6A)有一个简单的感光器官,称为眼珠(图6D),它包含少量纤毛光感受器,周围有一个大的色素细胞。有人认为这个眼胞是一个祖先成对的眼胞的残余物(48)。幼虫没有做什么,只是游到水底,把头埋在岩石上,溶解自己的神经系统,把自己变成一个水花。
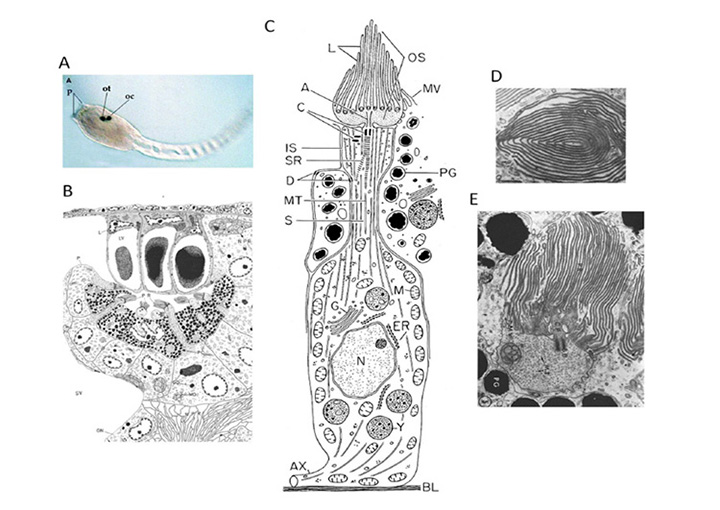
感光细胞。两种海鞘的纤毛感光器见图6B-E(来自(49,50))。总共有十几个左右的感光细胞(图6B中约有七个被切片)从单个色素细胞中伸出,它们的外层位于由三个晶状体囊泡(LV)细胞形成的晶状体下面。一个图示Ciona感光细胞如图6C所示。一个大的体细胞区域在它的基部产生轴突(AX),而从它的上端,一个突起穿透着色细胞并产生轴突(A),轴突通过它的片层(L)扩展外部段(OS)。Aplidium,得到了横向(图6D)和纵向(图6E)相对于外节段的轴,并显示了相当“花瓣状”的薄片同心排列。上述两项研究都报道了微绒毛与片层相互交错,尽管Eakin和Kuda(50)认为微绒毛起源于色素细胞(图6C),而Barnes(49)则报道微绒毛起源于光感受器的内段(图6E)。
电生理学。海鞘睫状体光感受器的电反应仅在一项研究中有记录Aplidium constellatum(51),据报道,“事实证明,从这些制剂中记录电活动极为困难,可能是因为靠近小动物表面的视网膜细胞往往在去除厚外衣时受损”。罕见的渗透给出了-5到-20 mV的静息电位,带有小振幅(2到7 mV)的超偏振光响应,伴随膜电导的降低。这些反应在性质上与脊椎动物视网膜光感受器的反应相似(振幅小可能是由损伤造成的),尽管没有足够的数据允许进行适当的比较。
八目鳗类鱼(Eptatretus物种)
现在有有力的证据表明盲鳗是七鳃鳗类祖先的后代,它们的许多形态特征是从更复杂的形式“退化”而来的。这种解释在这里当然是被接受的,但是,尽管如此,下面将提出的观点是盲鳗的眼睛提供了一扇窗口,以了解脊椎动物眼睛进化的早期过渡形式,正如Lamb等人(3)之前所建议的那样。
尽管盲鳗的系统发育位置一直存在争议,但现在有有力的证据表明盲鳗与七鳃鳗是一个进化支(52),如图1所示。盲鳗的身体平面是所有脊椎动物中最简单的(图7A, B)。它们生活在大多数大陆周围的海洋中,通常在很深的地方(很多情况下是200米或更深),在那里它们以掉落的尸体为食;最近人们发现它们也具有掠食性(53)。
在行为上,盲鳗似乎是瞎的。然而,在圈养环境中,暴露在明亮的光线下,经过长时间的延迟(10秒或更长时间)后,会开始游泳,有时还会尝试钻洞。除去动物的眼睛后,Newth & Ross(54岁)报告说,时间延迟没有变化,而Kobayashi(55岁)报告说,时间几乎翻了一番,从10秒左右增加到20秒。在这两种情况下,盲鳗的眼睛似乎对快速光敏行为没有什么调节作用。
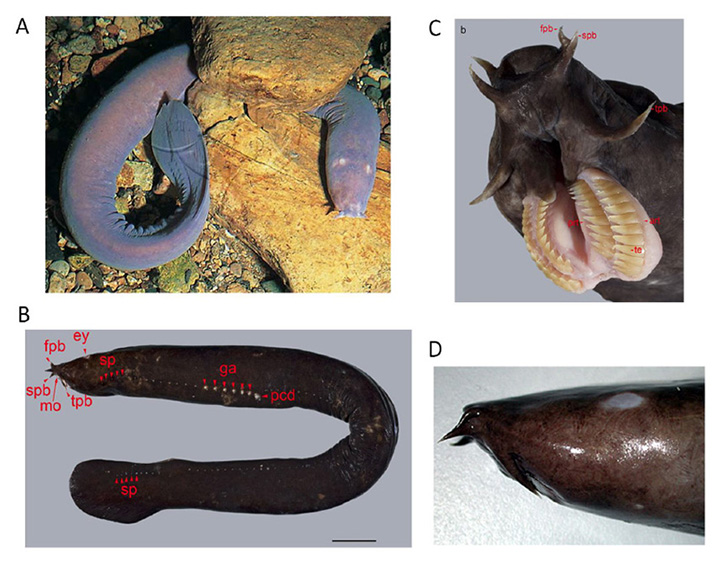
盲鳗能够从一系列侧孔中分泌出一种强力黏液(图7B),这种黏液是一种高效的防御机制。壮观的电影(补充视频S1,补充视频S2)显示,在捕食者咬盲鳗的那一刻,粘液迅速释放到它的嘴里,伴随着几乎瞬间的呕吐反应和捕食者的撤退。在拍摄到的14次攻击中,没有一次成功,每一次盲鳗都继续游泳,就像什么都没有发生一样。这种成功的防御机制的进化可能使盲鳗在它们的生态位中存活了数亿年,几乎没有改变。
八目鳗类鱼的眼睛.Locket和Jorgensen(56)对盲鳗的眼睛进行了全面的综述,盲鳗的眼睛在19世纪晚期首次被研究,然后在20世纪晚期使用电子显微镜。以下是对Eptatretus物种主要基于Holmberg(57,58)、Fernholm & Holmberg(59)和Locket & Jorgensen(56)的报告。
盲鳗眼的一般特征.在眼睛的头部位置,盲鳗只有一块半透明(几乎透明)的皮肤(图7D)。在这片半透明的皮肤下面是盲鳗的眼睛(图8A,图8B)。这个器官没有眼外肌,没有晶状体,也没有虹膜,而是嵌在脂肪中,一根非常纤细的视神经穿过脂肪。眼睛的大小因人而异,但通常在直径1 - 1.5毫米左右,但很少是球形的。巩膜/角膜不分为单独的不透明和透明区域,而是相当均匀的半透明。即使动物的皮肤可能是深色的,但在眼睛中,无论是巩膜、脉络膜或视网膜上皮中都没有发现色素沉着。巩膜内衬是一层纤细的毛细血管,大概可以与下颌脊椎动物的脉络膜血管系统相媲美。在没有晶状体的情况下,大部分腔体都被玻璃体填充。视神经很薄,只有几千个无髓神经轴突。 Neither hagfish nor lampreys exhibit myelin (60, 61), and these axons project predominantly to the hypothalamus (62, 63), just as their likely homologs, the melanopsin-expressing ipRGCs, do in mammals (64).
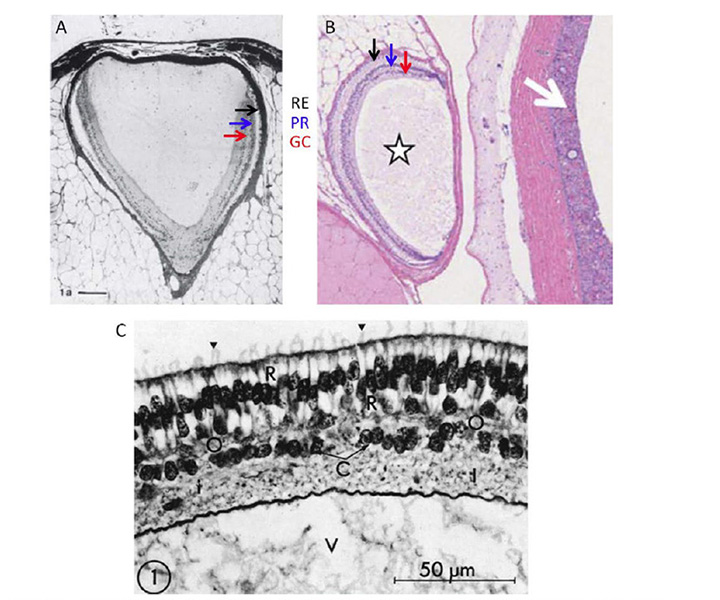
盲鳗的视网膜大致呈杯状,占地球面积的一半到三分之二,但脉络膜裂隙通常是开放的(65)。颌骨脊椎动物的视网膜由两个相对的层组成,神经视网膜和视网膜上皮,而盲鳗的视网膜上皮层是无色的。一些作者注意到,神经视网膜和视网膜上皮似乎经常被间隙隔开,但这有可能是一种固定伪物(可能是由类似海水的高强韧细胞外介质的稀释引起的)。在视网膜的外周边缘,内层减少为单层细胞,与外层细胞相连,但没有延伸到睫状体或虹膜。
视网膜神经.盲鳗的神经视网膜(图8B)比七鳃鳗或下颌脊椎动物的神经视网膜更简单,只有两层体体,包括感光细胞和投射神经元。虽然Locket & Jorgensen(56)报道了一些类似Landolt club的纤毛结构,但没有作者报道过可识别的水平细胞或双极细胞。盲鳗视网膜的神经排列与非哺乳脊椎动物的松果体器官非常相似(见第3节),有趣的是,盲鳗没有松果体。据推测盲鳗的光感受器与投射神经元(神经节细胞)有直接的突触接触,尽管到目前为止光感受器突触上的突触后细胞的身份还没有确定。视网膜有明显的内外膜包围。
光感受器的形态。盲鳗光感受器的形态Eptatretus stoutii如图9(57)所示。图9A的示意图显示了大致的排列,大致为圆柱形的受体细胞(R)被胶质细胞(G)包围。与下颌骨脊椎动物和七鳃鳗的情况相反,内段对应的区域位于外膜的玻璃状。它包含一个与椭球体相对应的区域,挤满了线粒体(图9C),纤毛从这里产生,但没有抛物面或肌样体的迹象。外段通过外限界膜突出进入心室/细胞外间隙(ES),在那里它与非色素上皮细胞(E)和从该细胞下来的细突接触。与其他脊椎动物的视网膜相比,其外节段在心室间隙中排列得非常稀疏(图8B和图9A)。
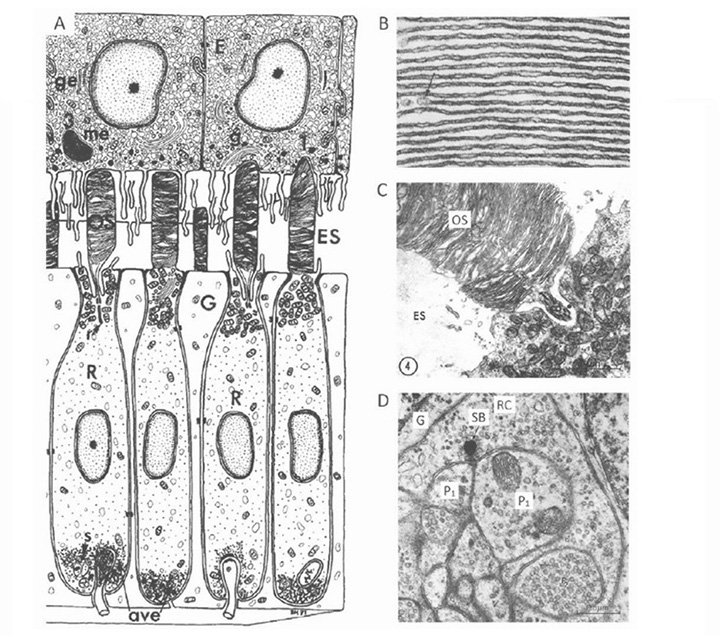
图9B和图9C的电子显微图显示了外层膜的层状排列。根据Holmberg(57)和Locket & Jorgensen(56)的研究,片层被质膜包围,尽管从显微照片上看不清楚。一些作者报告片层相当规则(图9B(57)),而另一些作者则不是;例:“这些碟片Eptatretus然而,它们并不是紧密地堆叠在一起,而是以一种松散且往往是无序的方式”(56)。纤毛具有经典的9+0双丝结构,但不寻常的是位于中心,在内外节的轴线上,因此外节片片大致对称地延伸在纤毛的两侧(图9C);这与脊椎动物锥体和杆状动物的情况相反,它们的纤毛位于外节段的边缘。
在其基部,受体细胞被突触触点内陷(图9A),突触区(图9D)的特征是突触囊泡围绕着“突触体”(SB),而不是传统的突触带。这种接触是二分体型的,而不是七鳃鳗和大多数下颌脊椎动物的光感受器中的三分体,尽管(如上所述)尚未有关于突触后元件的身份的报道。然而,由于没有其他细胞类型的报道,因此推测光感受器与投射神经元之间一定有直接的突触接触。
电生理学。虽然Kobayashi(55)报道了盲鳗的ERG记录,但还没有从盲鳗的光感受器中获得单细胞记录Myxine garmani但后来被Fernholm & Holmberg(59)报道Eptatretus burgeri.使用被切除的眼睛,在眼睛表面的灯芯电极和眼睛放置的湿布之间进行记录。微弱的闪光引起一种典型的先正后负形状的缓慢反应,这种复杂反应的振幅在~10勒克斯的相对较低的强度下饱和。对于明亮的闪光,在这个复杂的反应之前会有一个小的缓慢的负向波。反应的光谱灵敏度在500 nm左右达到最大值,提示存在视紫红质。
尽管Kobayashi将复杂的正-负波解释为类似于脊椎动物眼睛的ERG的b波,但有一些显著的特性表明了不同的解释。首先,这种反应表现出一个相对较长的潜伏期,大约350 ms,然后在大约200 ms内迅速上升到它的正峰值。其次,当闪光强度进一步增加或闪光持续时间从3 ms到1 s变化时,这种复杂反应的形式完全不变。第三,反应表现出较长的不应期,因此在10勒克斯闪光后需要约4秒的间隔,才能从第二个相同的闪光中引起任何反应。第四,昏暗的适应光可以完全消除这种反应,然后在随后的黑暗适应中,这种反应又突然出现。
综合这些特征,我提出了另一种解释,即先积极后消极的反应实际上反映了投射神经元(神经节细胞)中非常缓慢的再生电位(“动作电位”)的同步激发。未来,通过对盲鳗视网膜细胞进行更全面的电生理记录,包括神经节细胞的细胞内记录,光感受器的吸液吸管记录,以及进一步的ERG记录,应该可以验证这一论断。
盲鳗的光感受器表现出一些杆状的特性。总之,光感受器Eptatretus物种表现出许多棒状的特性:例如,据报道外段片层为盘状,内段缺乏抛物面或肌状,电响应在500 nm左右显示出峰值灵敏度。然而,仍然没有证据表明这些细胞是否能对单个光子做出可靠的反应。
七鳃鳗松果体(Petromyzon绿ammocoete)
就像侧视眼的视网膜一样,松果体是间脑的外展,尽管它在背中线上向上出现(见第13节)。在非哺乳脊椎动物中,松果体中含有光敏纤毛光感受器,所有的超微结构研究都表明,只有三种主要细胞类型存在:光感受器、投射神经元(神经节细胞)和胶质细胞。光感受器在神经节细胞上形成带状突触,神经节细胞将轴突发送到下丘脑。
海七鳃鳗幼虫松果体中的光感受器,Petromyzon绿Pu & Dowling(66)使用光镜和电子显微镜以及细胞内电生理记录进行了研究,以下是基于他们的报告。图10显示了松果体光感受器及其在松果体中的组织。图10A的示意图描绘了器官的总体布局,而图10B的示意图则说明了感光细胞及其突触接触的详细特征。
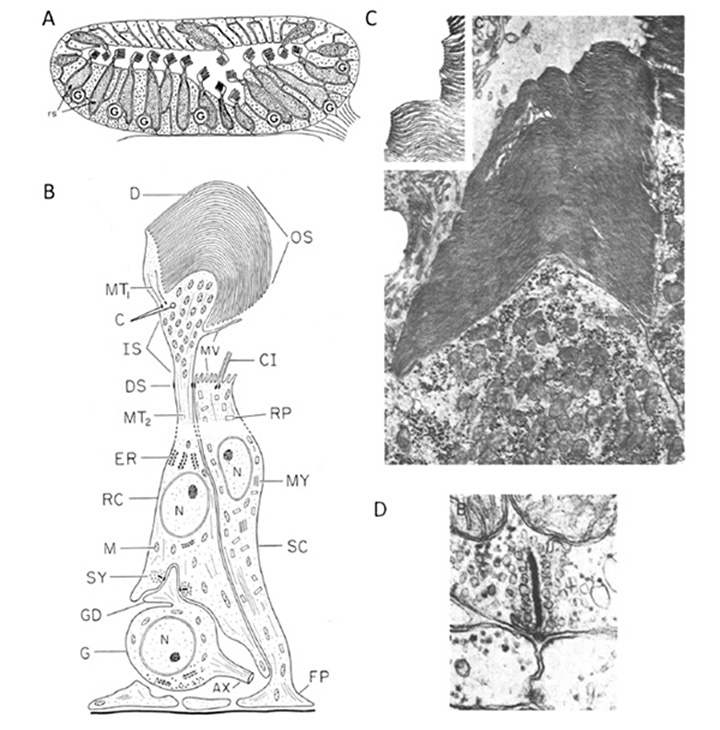
光感受器的形态。如图10B(67)所示,光感受器细胞的大部分长度近似为圆柱形,直径接近10µm,但外段通常较宽,宽度为10 - 25µm。外节段含有大量的片层,其数量可超过100个(图10c),这与Nilsson(13)认为松果体膜“膜层堆积非常有限”的观点形成鲜明对比。片片很少是平的,但通常有点弯曲,据报道,它们在颚口视网膜的锥体和杆状外节的堆叠有点不规律。外段膜类似于锥体膜,与质膜连续(图10C)。
与外节段一样,突出的内节段突出到腔内。在细胞基部,突触与神经节细胞接触,主要是在扁平的带状突触上(图10B,图10D),排列为成对(图10D)或“单子”;三联征从未被发现,反馈突触也未被发现(66)。
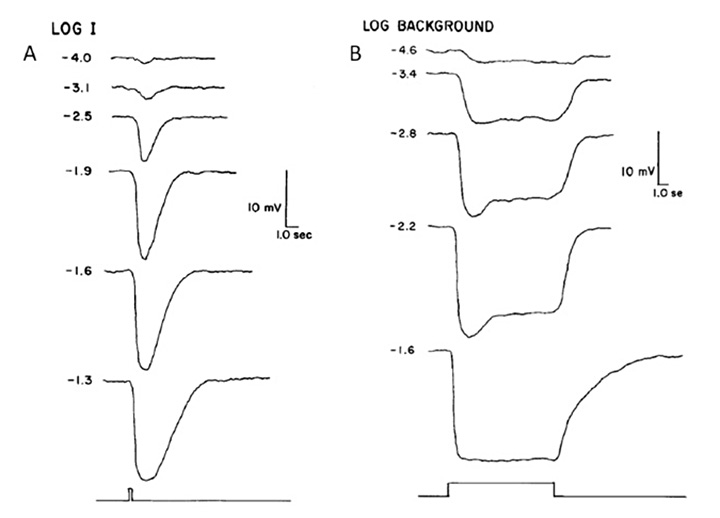
对光的电响应.Pu & Dowling(66)从这些松果体光感受器中记录了细胞内电压,对闪光和光阶的响应如图11所示。这种反应总是一种缓慢的渐变超极化,与脊椎动物锥体或杆状体记录的超极化大致相似,但没有对明亮刺激从最初的峰值迅速放松到平台期的特征。其光谱灵敏度在545 nm左右达到峰值。微弱闪光达到峰值的时间约为1 s(图11A),比室温下的视锥光点要慢得多,但与视杆光点相似。闪光的响应-强度关系遵循双曲线饱和,I / (I+σ)。在长时间的光照下,响应缓慢下降,除了在最高强度时,它保持饱和(图11B)。对增量闪光的响应是脱敏的,大致根据韦伯定律,尽管大部分脱敏是响应压缩的结果,只有大约1.5 log单位是由于σ的缩放。当稳定光停止时,这种反应立即开始恢复,就像锥细胞一样。在强烈的“漂白”暴露后,灵敏度在4分钟内完全恢复,再次类似于球果而不是杆状细胞。
Kusmic等人(68)在鳟鱼松果体光感受器中获得了广泛的可比性结果,并进一步表明分子机制似乎与视网膜光感受器相似。电压钳实验表明,光响应伴随着膜电导的降低(如视网膜光感受器);然而,明亮的闪光只降低了约10%的总电导,这可能解释了Morita等人(69)关于光诱导电导增加的相反报告。此外,应用磷酸二酯酶抑制剂IBMX导致光反应的大小增加,就像视网膜光感受器一样,并与光诱导的cGMP浓度下降一致。此外,在固定时间(峰值之前)测量的响应-强度关系显示出指数饱和,正如经典脊椎动物光转导级联所预期的那样(70)。
Kusmic等人(68)报道的光适应行为与Pu和Dowling(66)报道的略有不同,因为对稳定光照的响应完全没有下降。此外,对于叠加测试闪光,在背景照明的存在下,达到峰值的时间并没有缩短。对于昏暗的测试闪光,在黑暗适应条件下达到峰值的时间约为1.5 s,在有背景的情况下保持不变;同样,对较亮的测试闪光(在黑暗中达到峰值的时间较短)的反应也没有加快。
从Pu和Dowling(66)报告的光谱灵敏度,以及Koyanagi等人(71)的原位杂交结果来看,上面记录的松果体光感受器中的视蛋白似乎是视紫红质(尽管含有维生素a)2),这些细胞可能来自器官的腹侧区。背侧区域的光感受器表达副视蛋白(71)并表现出对紫外线敏感的超极化(71,72)。
尽管上述松果体光感受器可能使用视紫红质作为视觉色素,但除了两个特性外,它们的电反应一般可以描述为类似“缓慢而不敏感的锥状反应”。首先,光适应没有响应加速。其次(可能是相关的),响应在明亮稳定的光线中饱和。相比之下,无论稳定的光线有多亮,下颌脊椎动物的视锥细胞总是设法将循环电流和细胞内电压恢复到一个操作点,使它们能够继续对增加的刺激作出反应(73)。
松果体神经节细胞反应。一个标准的“光度OFF”神经节细胞对中等亮度照明的典型反应如图12(69)所示,并与光感受器的反应进行示意图比较。对于次饱和强度,神经节细胞中的分级反应形式与光感受器中的反应形式非常相似。在其他的实验中已经表明,持续的曝光降低射击速率与强度的对数成正比,在高达8个对数单位(74)的范围内。
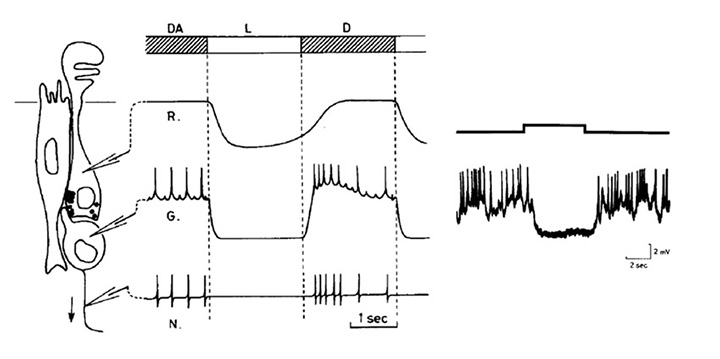
七鳃鳗侧眼
的眼睛。成年七鳃鳗的侧眼(图13B、图13C)与颌鱼的侧眼惊人地相似。这是一个照相机式的眼睛,有一个晶状体,一个虹膜和一组六块眼外肌肉。这些眼外肌肉在一定程度上与下颚脊椎动物的肌肉同源(75),有趣的是,在一种皮鳞鱼化石中记录了中间排列的肌肉(77,77)。皮鳞鱼是一种带甲的鱼,在七鳃鳗的祖先分化后从我们的谱系中分化出来。

视网膜。如图14所示,银七鳃鳗(一种北半球物种)的视网膜与颌口目动物的视网膜非常相似,并包含传统的五类神经元(光感受器、水平、双极、无分泌和神经节细胞)以及Müller胶质细胞。细胞核分布在三个主要的核层中,还有两个丛状层,尽管七鳃鳗和颌口视网膜的一个区别是神经节细胞层和内丛状层的明显“翻转”——因此,视网膜神经节细胞及其纤维的大部分位于七鳃鳗的内丛状层(比较图14A和B)(75)。Fritzsch提出,七鳃鳗视网膜(以及某些大脑区域)的这种排列是基础的,并由此衍生出颌口动物视网膜层的翻转(77,79)。
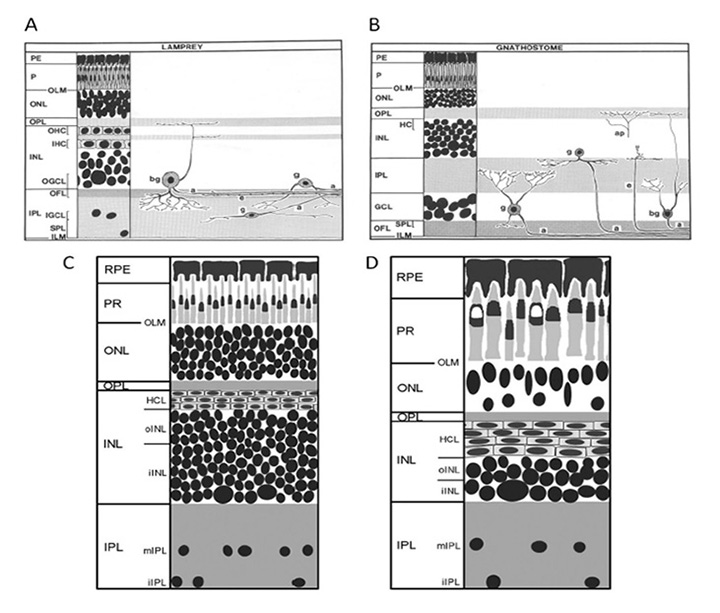
图14C、D所示为南半球七鳃鳗的视网膜示意图,Geotria南极光的“下游”和“上游”迁移阶段;下游阶段只是后变质阶段,当幼鱼洄游到海里时,而在上游阶段,成年的成虫洄游回上游,在它们孵化的地方附近产卵。视网膜的g .南极光与其他七鳃鳗大致相似。图14C、D显示了在海洋间相期间发生的光感受器尺寸的大幅增加。
视网膜内氨基酸、神经递质和钙结合蛋白的免疫反应性分布g .南极光最近Nivison-Smith等人(80)对其进行了研究,并表明其与颌骨脊椎动物的发现大致相似。小有机阳离子胍丁胺的实验结果与阳离子进入光感受器和水平细胞的结果一致,这与颌骨脊椎动物视网膜的情况非常相似。
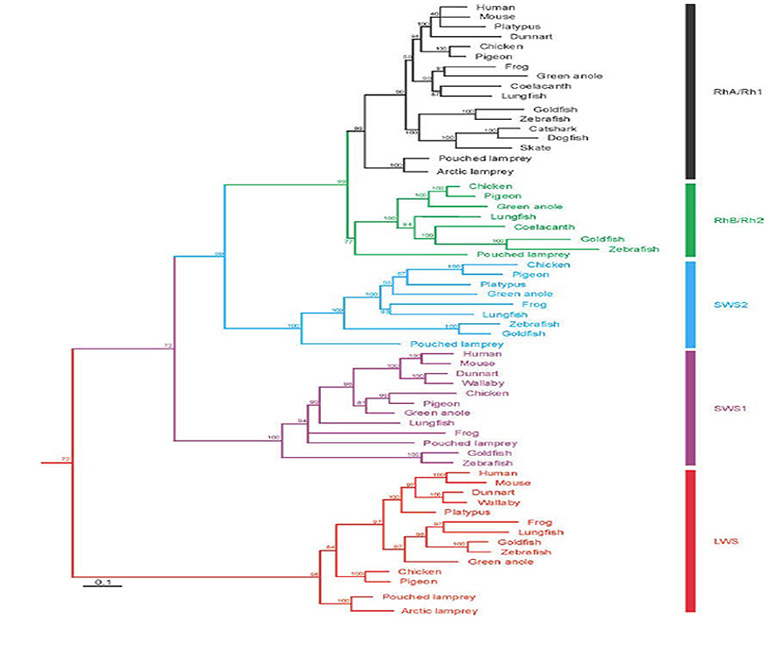
视蛋白和光感受器的种类。七鳃鳗视蛋白分为五类,似乎与下颌脊椎动物的视蛋白同源(或近似同源);因此,南半球的七鳃鳗Geotria南极光明显具有LWS, SWS1和SWS2视蛋白,其其余两种视蛋白,RhA和RhB,可能分别属于Rh1和Rh2家族(81,82)。此外,该物种拥有五种不同的感光细胞(83)。虽然视蛋白类在光感受器类中的表达分布尚未得到明确的确定,但间接证据表明图15B所示的分布(Shaun P Collin,个人交流)。与之相反的是g .南极光其他种类的七鳃鳗失去了不同数量的视蛋白和感光细胞;因此,南半球的另一个物种Mordacia mordax只有一类视蛋白和一类光感受器,而北半球的物种一般有两类视蛋白(Rh1和LWS)和两类光感受器。
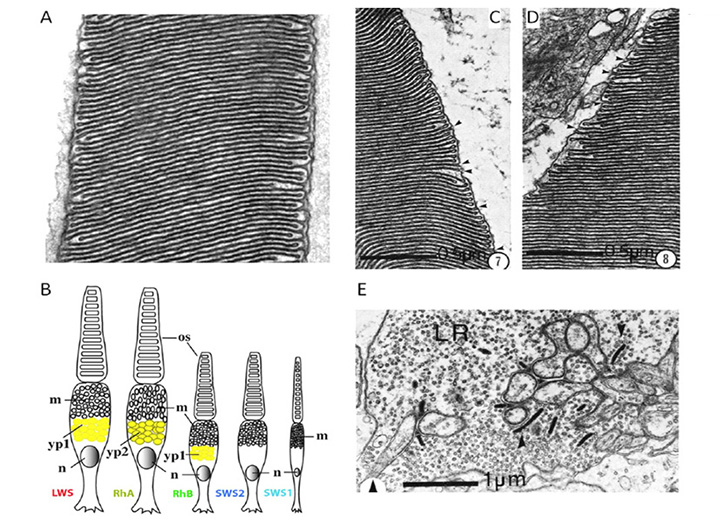
光感受器的形态。在北半球的七鳃鳗中,这两种类型的光感受器根据其内部节段的长度被称为“长”和“短”;有些令人困惑的是,它们的外部部分与此相反。因此,“长”细胞在与视网膜色素上皮密切接触的远端层有短的(~7 μ m长)圆锥形外段,而“短”细胞(其数量比长细胞多出3:1)在近端层有较长的(~25 μ m长)圆柱形外段,且只有顶端达到RPE。
七鳃鳗视网膜感光器的超微结构Petromyzon绿Dickson & Graves(84)进行了检查,他们报告说这两类细胞(短细胞和长细胞)表现出锥形而不是杆状的形态。因此,在两种细胞类型中,外段膜似乎与质膜是连续的。尽管发现有小群的囊/盘被质膜包围,但其外部经常有开口,如图15C, d所示,带有标记氨基酸的放射自显影显示,新合成的蛋白质均匀分布在外段,如球果中所示。此外,外部节段从未显示出切口(表面膜的纵向深嵌层,它将颌骨脊椎动物的杆状椎间盘分成小叶)。最后,突触末梢(图15E)被报道类似锥椎弓根而不是杆状球体。因此,在所有用于区分颌骨脊椎动物锥体和杆体的常规标准上,的短和长受体p .绿都属于视锥细胞。
在南半球的七鳃鳗中也得到了大致类似的结果,g .南极光,其中超微结构检查导致提出所有五类光感受器都是锥形的(83,85)。在这五类光感受器中,每一类的外段膜都与细胞外基质连续(图15A),突触末端包含1到5个突触带。此外,至少有三种细胞类在内段含有过滤色素(图15B)。
phototransduction的蛋白质。如第8节所述,视蛋白、转导蛋白、PDE催化和调节亚基的异构体分布已经确定p .绿Muradov等人(86,87)。长受体表达一种LWS视蛋白,一种转导α亚基GαL,可能是祖先的,一种常见的PDE6,似乎是祖先的,和一种PDE γ亚基,与颌口锥亚型进化。短受体表达一种Rh1视紫红质,一种杆状转导素α亚基GαS,但仍然保留锥形特征,常见的PDE6似乎祖先,和PDE γ亚基与牙突杆亚型进化。
七鳃鳗基因组。Smith等人(88)最近报道了七鳃鳗的第一个全基因组序列和组装p .绿.分析表明,在脊椎动物谱系的基础附近发生的“2R”两轮全基因组复制早在七鳃鳗和颌口鱼谱系的分化之前就已经发生了(即在图1中的5之前)。
电生理学。Govardovskii & Lychakov(89)研究了光感受器的长、短ERG响应特性七鳃鳗丁。在暗斑强度下,他们观察到由517纳米色素(已知存在于短细胞中)驱动的b波,尽管这种ERG反应不像在青蛙视网膜中看到的那样敏感。在光强下,a波是两类光谱信号的组合,两组光谱响应均表现出光适应性,不能饱和。因此,虽然长细胞和短细胞都具有锥状形态,但短细胞的电生理行为有点像杆状细胞,但两类细胞都表现出锥状适应。
“杆状体”存在于远古的脊椎动物中吗?如上所述,七鳃鳗'视紫红质' RhA/Rh1与下颌脊椎动物的视紫红质Rh1密切同源(82,83,90)。Pisani等(82)对序列分析的解释表明,七鳃鳗和下颌脊椎动物的共同祖先拥有Rh1基因。他们继续说:“Rh1在agnathans中的功能尚不清楚,但假设它在脊椎动物生祖中的功能与它在大多数脊椎动物中的scotopic功能没有显著不同,这意味着光和scotopic视觉都在茎脊椎动物谱系中进化,并且在大约522-518 Ma的寒武纪就已经存在了。”然而,目前还没有足够的证据证明这一点,因为Rh1视蛋白是否在“真棒”中表达,或者Rh1是否介导了“暗斑视觉”。根据目前的证据,完全有可能祖先的含有rh1的光感受器只是作为一个“慢敏锥”,而不是作为一个真正的能可靠地检测单个光子的棒,这是为了使视觉系统达到最终的scotopic性能所需要的,而这种性能是在下颌脊椎动物中所达到的。beplay体育公司从这些证据中可以肯定的是,有颌和无颌脊椎动物的共同祖先极有可能拥有一种杆状视蛋白;但这还不足以推论出它有暗斑视力。
七鳃鳗光感受器特征综述。北半球七鳃鳗有长短光感受器,长细胞具有颌骨脊椎动物视锥细胞的所有特征。短细胞具有视锥细胞的大部分特征,尽管它们表达视紫红质以及某种杆状转导素α亚基和杆状PDE调节亚基;光转导的其他组成部分还有待分类。短细胞有很高的灵敏度,尽管到目前为止还没有证据表明它们可以可靠地显示单个光子的撞击。在南半球七鳃鳗中,有五类光感受器,它们似乎都具有视锥细胞的大部分特征。其中一种表达的是视紫红质,但目前还没有足够的证据表明它是否具有杆状的功能特性。
下颚脊椎动物的侧眼
所有下颌脊椎动物的眼睛在整体布局和特征上都非常相似。视网膜有相同的细胞种类,并且以基本相同的方式组织。此外,除了大小不同外,所有颌口动物的锥状感光器(以及杆状感光器)在结构上非常相似,它们对光的电响应也非常相似。鉴于从第一种有颌的脊椎动物出现以来已经过去了4亿多年,令人印象深刻的是,眼睛的基本结构以及光感受器的结构和反应特性几乎没有发生变化。
下颌脊椎动物的锥状和杆状光感受器的结构和响应,以及光转导过程在其他地方有全面的讨论(参见Webvision的其他部分;埃布雷和库塔洛斯(91岁);等等),这里不再重复。为了完整起见,图16展示了哺乳动物光感受器的主要超微结构特征,以便与其他脊索动物的光感受器进行比较。锥和杆的功能特性将在第7节进行对比,而转导级联的分子成分将在第8节进行回顾。
4脊索动物眼的分级:视网膜和感光器
从第3节的描述中,我们可以看出视网膜和睫状光感受器类型的分级,现在将对其进行描述。同样地,人们发现视蛋白(c -视蛋白)的性质的分级,这些将在第6节中介绍。
脊索视网膜的分类
现存脊索动物的光敏神经组织可分为三类:
一个没有“视网膜”)。“原始的”脊索动物(头脊索动物和被囊动物)没有视网膜(通常使用这个术语),而是分别在前眼和眼珠中有少量与色素细胞相关的睫状体光感受器。在囊胞中,纤毛光感受器每一个都产生一个轴突,但其突触接触尚未确定。在头脊索前眼中,输出轴突似乎来自第二排细胞,而不是睫状光感受器本身。
b)两层视网膜。盲鳗的侧“眼”和非哺乳动物下颌脊椎动物的背侧感光器官(松果体/松果体旁/顶叶)呈现出两层视网膜,纤毛光感受器通过带状突触与投射神经元(神经节细胞)进行接触。光感受器和投射神经元嵌入在胶质细胞中,但还没有发现其他种类的神经元(虽然不能排除它们的存在)。
c)三层视网膜。七鳃鳗和下颌脊椎动物的侧眼有三层视网膜,在睫状体光感受器和视网膜神经节细胞之间有双极细胞,另外还有两类神经元,以水平细胞和无分泌细胞的形式存在。
脊索睫状体光感受器形态类型的分级
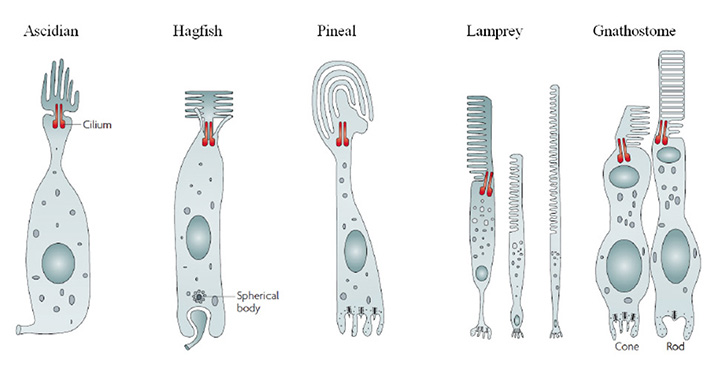
在形态学的基础上,第3节中描述的脊索视网膜的睫状体光感受器细胞可以描述为图17所示的示意图。在所有情况下,感光的外部部分是由膜表面的大量膨胀形成的,以片层的形式产生纤毛,其特征是9+0双丝。这一外层膜充满了高浓度的视觉色素,以及较低水平的其他光转导蛋白。细胞体在大多数情况下大致呈圆柱形,位于细胞核的上方和下方。突触输出发生在细胞末端相对于外节段(即“基部”)的地方,有时外节段与体细胞被中间的“轴突”节隔开。这些特征至少可以分为以下五组:
(我)Ascidian-style。外节片的排列很像花瓣,大致纵向地围绕位于中心位置的连接纤毛。轴突离开了细胞的基部,但它的突触接触点尚不清楚。
(2) Hagfish-style。外节片向外伸展到连接纤毛的两侧,纤毛位于中央。突触输出发生在基底内陷处,突触囊泡围绕“球形体”排列,而不是丝带状。
(3) Pineal-style。外节片片非常宽,常呈曲线状。一般报道,片层构成锥状囊,而不是杆状盘。基底内陷突触呈突触带,与神经节细胞形成双(有时为单)接触。
(iv)锥。外节片层(囊)排列高度规则,其膜与内节的质膜形成连续统一体。哺乳动物的外节大致呈圆柱形,而非哺乳动物的脊椎动物的外节则趋向于圆锥形。内段可以包含各种特化,包括椭球体(有线粒体,有时还有光谱过滤的油滴或囊泡),收缩肌体和抛物面。
(v)棒。在形态学上,杆状细胞与锥状细胞非常相似,但有一个显著的例外是与外节膜的拓扑有关。在外段合成过程中,会产生锥状囊,但这些囊与相邻的囊在边缘周围形成密封,形成与质膜分离的盘,结果是狭窄的椎间盘内空间与细胞外空间隔离。
渐变为眼睛进化过程中的渐变提供了证据。上面列出的视网膜和光感受器类型的分级提供了有力的证据,证明脊椎动物的眼睛不是“突然出现的”,反而支持了我们的眼睛是通过无数微小变化进化而来的观点。这一结论(到目前为止)主要基于睫状体光感受器的形态,从视蛋白的分级分析(第5节和第6节),从转导级联的分子遗传学分析(第8节),从胚胎眼发育的研究(第13节和第14节)得到了极大的强化。
5Pre-vertebrate脊索动物C-opsins
由于最近的工作,已经有可能跟踪使原始脊索c视蛋白(表现出许多与r视蛋白相同的特性)进化为现代锥视蛋白和杆状视蛋白的直接前体的变化。
热稳定性和光逆转
锥状和杆状c视蛋白与它们的r视蛋白表亲(包括黑视蛋白)有很大区别的两个方面与光激活all-的热稳定性有关反式“后视视紫红质”状态及其光逆转能力(图18)。
在r -视蛋白中,光激活的后视视紫红质是热稳定的(图18右),其半衰期通常为数小时甚至数天。这种活性形式被抑制素分子的结合迅速灭活,但它在其全-中保持稳定反式配置。在大多数情况下,这个后视紫红质在光谱的可见区域有它的峰值吸收,这表明,在它的酶活性构型中,所有-的席夫碱键反式视黄醛是质子化了的。此外,在吸收了另一个光子之后,这个稳定的all-反式后视紫红质(即使被抑制素结合)也可以通过光逆转回到它的11-独联体视紫红质形式。事实上,在大多数实际用途中,这种光逆转是许多微绒毛(横纹肌)光感受器中视觉色素再生的唯一短期机制。
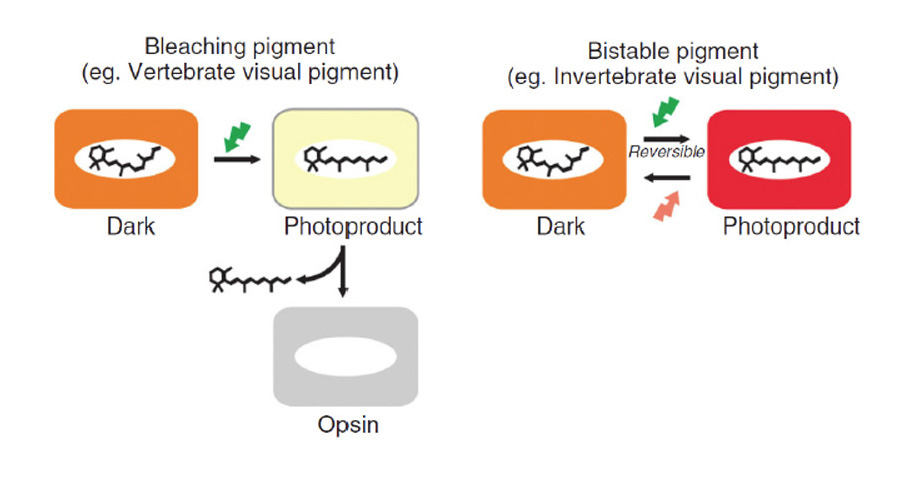
相比之下,锥体视蛋白和杆状视蛋白的光激活后视紫红质II状态是热不稳定的(图18,左),锥体视蛋白的半衰期较短(秒),而视紫红质的半衰期稍长(分钟)(值见Imai等人(92)的表2)。快速失活是由于阻滞素结合的结果,通过快速磷酸化实现。活性元II在UV (~380 nm)中吸收,因为席夫碱现在没有质子化。尽管质子化的meta I中间体可以经历光逆转到11-cis构型,但活性meta II态不能经历这种光逆转,即使它吸收了一个蓝色/UV光子(见Ritter et al .(93)的图7),而这种能力显然是伴随激活而来的内部分子重排的结果。
脊索c视蛋白的系统发育和性质分级
c视蛋白是一个大家族,包括刺胞动物和原口动物,以及后口动物(包括脊索动物和脊椎动物)。除了众所周知的视网膜锥视蛋白和视蛋白杆视蛋白,脊椎动物还表达其他几种c视蛋白,包括:Opn3(脑视蛋白/ TMT视蛋白),parietopsin, parapinopsin, VA(古老脊椎动物)和视蛋白。从它们的分子系统发育,以及从已研究的五组视蛋白的功能特性的分级(现在将进行描述)来看,这些c -视蛋白反映了锥视蛋白和杆状视蛋白出现之前的进化顺序。
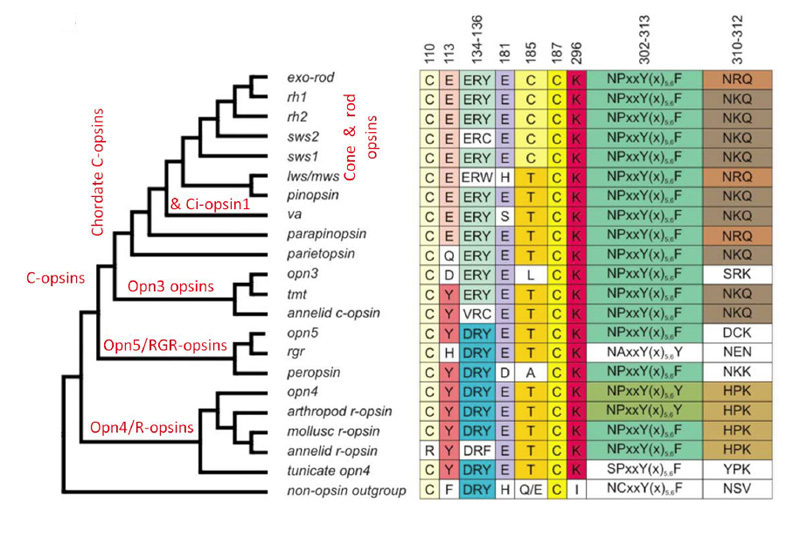
图19展示了视蛋白的简化分子系统发育,以及在一些重要位置的氨基酸残基的列表(94)。在左边的分支图中,请注意,线的长度并不表示进化距离,因此分支点的水平位置并不表示发散的时间。在右边的表格中(以及整篇文章),注意残基是按照牛视紫红质序列编号的。
如图19所示,脊椎动物c -视蛋白从最古老的到最近的顺序为:Opn3(脑视蛋白),顶视蛋白,副视视蛋白,VA视蛋白,视视蛋白,视锥视蛋白,视紫红质。对于每一种c视蛋白,其功能分子特性简要描述如下,并在表2中总结;这些信息大部分来自于最近对重组蛋白的研究。
在评估这些c -视蛋白的相对性质时,将与“典型的”r -视蛋白进行比较,显示两种稳定状态之间的光可逆转变。如上所述,脊椎动物视蛋白锥和视蛋白杆不显示光逆转,而是释放它们的所有-反式类维生素a。另一方面,它们对g蛋白的激活更强。现在我们将看到其他脊椎动物的c -视蛋白表现出介于这两者之间的特性。
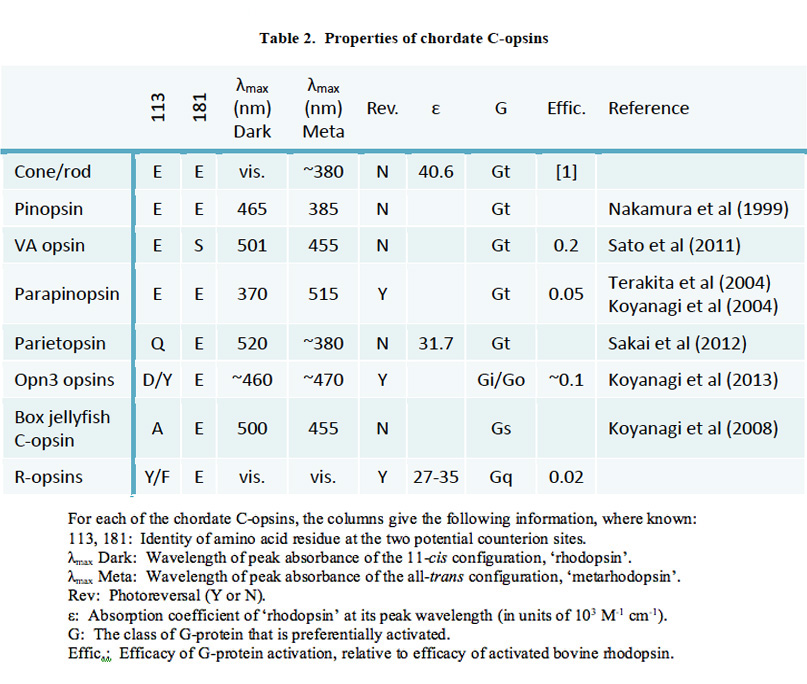
Opn3。Opn3视蛋白组包括哺乳动物Opn3视蛋白(以前称为脑视蛋白或泛视蛋白)、TMT(硬骨鱼多组织)视蛋白、昆虫翼视蛋白和环动物c视蛋白。该基团在c -视蛋白的基部附近演化,其序列与r -视蛋白的序列非常相似。对大多数基团的成员来说,质子化席夫碱的反离子的两个潜在位点(113和181)为r -视蛋白形式,Y113和E181,表明E181作为反离子;然而,在哺乳动物中,Opn3含有D113。哺乳动物的Opn3在脑深部分散的细胞中表达。
Opn3视蛋白的分子特性直到最近才通过在培养细胞中的表达进行研究(95)。河豚鱼TMT视蛋白与11-cis视网膜结合,形成一种吸收蓝色(约460 nm)的视觉色素;这种色素是双稳定的,被异构化为轻微红移的全-反式由短波长光形成,并异构化回11-独联体由长波长光形成。光激活型可激活Gi和Go,但其活性仅为激活型视紫红质的10%左右,而不能激活Gq、Gs和Gt(转导蛋白)。在蚊子的Opn3同系物中也发现了类似的结果,尽管它的活性更高。尽管哺乳动物Opn3本身尚未在培养细胞中成功表达,但Koyanagi等人(95)认为它很可能表现出类似的特性。
Parietopsin。壁蛋白最初是在蜥蜴顶眼的光感受器中发现的。Sakai等人(96)最近对其功能性质进行了研究,他们发现Schiff碱反离子位于181的“无脊椎”位置,与残基Q113和E181一致。表达的色素在520 nm处有吸收峰,其光敏度和摩尔消光系数略低于锥视蛋白和杆状视蛋白,这是无脊椎视蛋白的通常特征。另一方面,它的光化学性质类似于锥状和杆状色素,形成的是腐烂的后视紫红质中间产物(I、II和III),而不是典型的无脊椎动物光可逆的后视紫红质。
Parapinopsin。旁棘视蛋白最初是在鲶鱼的棘旁器官中发现的。它随后在七鳃鳗松果体中被发现,被证明是一种紫外线敏感的双稳态视蛋白(97)。Koyanagi等(71)发现鳞视蛋白旁只在七鳃鳗松果体和松果体旁器官的远端纤毛光感受器中表达,并对其功能特性进行了研究。他们发现,副视蛋白的照射引起了传统的11-独联体所有- - -反式生色团的异构化,产生光谱峰在515 nm处的产物。然而,与紫外线敏感(SWS1)锥视蛋白的情况相反,这种后视视蛋白被发现是稳定的,并在吸收后续光子时表现出光逆转;事实上,通过UV和橙光的连续照射,副视蛋白和它的元态之间可以反复相互转化。表达副视蛋白的松果体光感受器的细胞内电记录显示,紫外线闪光引起缓慢的超极化反应,与脊椎动物的光转导级联一致。活化的副紫视蛋白活化g蛋白的效果比活化的视紫红质低约20倍(97)。
VA / VAL视蛋白。VA视蛋白首先在鲑鱼的视网膜中被发现,然后很快在其他硬骨鱼的视网膜中被发现,在其他硬骨鱼的视网膜中也发现了较长的剪接变体(VAL),然后在其他脊椎动物中也发现了。Sato等人(98)最近研究了VAL视蛋白的功能性质。像脊椎动物视视蛋白一样,VA/VAL在正常位置有E113,用于反位到静息状态,但独特的是,原始反位位置的位置是S181。Sato等(98)发现光活化引起独联体来反式但活化的中间产物在可见光区吸收,在455 nm处,表明席夫碱仍然质子化。虽然这种中间体吸收可见光,但它不表现出光逆转(脊椎动物视蛋白)。这种中间体可以激活Gi g蛋白,尽管其功效比激活的视紫红质低约5倍。因此,他们表明VA/VAL视蛋白的一些特性介于副视蛋白和视蛋白之间(见下文)。
Pinopsin。Pinopsin首次从鸡的松果体中克隆出来,并被发现与脊椎动物视蛋白的系统发育树底部附近的LWS锥视蛋白(99)发生演化。然而,目前还没有明确的证据表明它是在哪里或如何从视蛋白中分化出来的,很有可能它是祖先的。Nakamura等人(100)对其功能特性进行了研究,发现其与脊椎动物视蛋白的功能特性非常相似。静止的黑色形态在465 nm吸收,在光谱的蓝色区域,类似于SWS锥视蛋白(如SWS2, Rh2)的吸收。光吸收通过一系列光产物(batho, lumi, meta I, meta II, meta III)触发其进展,类似于激活视紫红质所触发的光产物。这些中间产物的寿命与锥视蛋白相当,除了元II状态,显示出更长的寿命更典型的视紫红质。
在脊索动物进化过程中c -视蛋白表现的改善
通过比较脊髓c视蛋白的序列、结构和功能特性,Akihisa Terakita和Yoshinori Shichida和他们的同事阐明了在进化过程中发生的一系列转变,这些转变赋予了脊髓c视蛋白更好的性能(Tsutsui & Shichida (101);冢本和寺田(102);Terakita等人(103))。这些变化包括反离子位点(对于质子化的希夫碱)从残基181转移到残基113,以及蛋白质的分子重排,在激活过程中提供了增强的螺旋6倾斜,从而导致了G-蛋白更强的激活。
反离子迁移。为了使静息视蛋白中的质子化希夫碱上的正电荷保持稳定,需要在附近有一个带负电荷的反离子。在大多数视蛋白(包括R-视蛋白、o-视蛋白、rgr -视蛋白和光异构酶)中,反离子位于181位(牛视紫红质编号中),由谷氨酸E181占据,而在锥视蛋白和杆视蛋白中,反离子位于113位,由谷氨酸E113占据。Terakita等人(97)发现了E181位置代表祖先位置的明确证据,并表明在脊索c视蛋白的进化过程中,反位位点已经迁移到113位置(图20)。
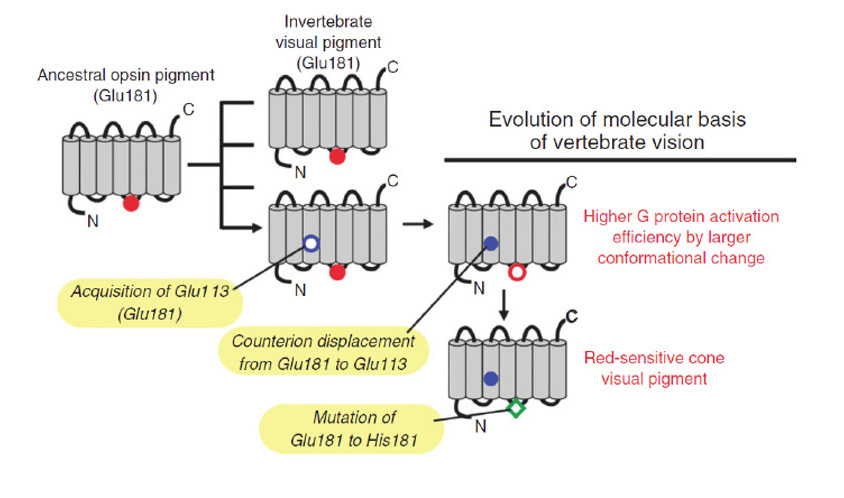
反离子的功能。除了在静息状态下稳定质子化希夫碱的作用外,锥体和杆状视蛋白中的E113反离子还有许多其他重要功能(Tsutsui & Shichida综述,(101))。也许最重要的是,它在激活过程中充当来自希夫碱的质子的受体(参见下面的分子机制描述)。最近,它被证明有两个额外的关键功能,在光吸收后使席夫碱键快速水解(这在视锥细胞中很重要)和同样地使11-快速结合独联体视网膜(104));这些作用将在下面描述,锥视蛋白和杆视蛋白的比较将在第6节中进行。反离子对静息视蛋白态的光谱调谐也有很强的影响。最后,它在降低静息视蛋白的本构活性方面也很重要;缺乏E113反离子的突变体在黑暗中通常表现出相当高的g蛋白激活率。因此,在脊椎动物视网膜视蛋白的进化过程中,获得这种反离子的替代位置被证明是非常重要的。
增强g蛋白活化的功效。研究发现(表2),光激活形式的锥视蛋白和杆状视蛋白对g蛋白的激活效果比r视蛋白的激活形式要大得多,而且在脊索c视蛋白中,这种特性是分级的。Tsukamoto等人(105)使用位点定向荧光标记技术来检测光激活过程中蛋白质的变化,他们发现激活过程中螺旋6相对于螺旋5的运动程度与视蛋白激活g蛋白能力的强度之间存在很强的相关性。
祖先视锥蛋白的前脊椎动物复制体。在脊索动物进化过程中出现的“改进的”c视蛋白,具有g蛋白激活的高效效果,并能快速释放all-反式视网膜,成为祖先视锥蛋白。此外,有强有力的证据表明,在脊椎动物谱系基础的“2R”全基因组复制之前,这种祖先锥视蛋白已经复制成为祖先的一对SWS和LWS锥视蛋白。但是,更方便的是以后考虑这些证据,在脊椎动物视视蛋白的第6节。
脊椎动物视紫红质活化的分子机制
牛视紫红质中后视紫红质状态激活的机制现在在分子水平上被理解,这在很大程度上是基于对激活形式的晶体结构的检查(106,107)。从根本上看,这种机制很可能适用于视蛋白,因此它出现在“2R”全基因组复制之前。图21提供了激活所涉及的事件的示意图(108)。在光子吸收的几微秒内,就会达到一种状态(meta I),在这种状态下,激发态保持在类视黄酮的扭曲中,蛋白质中几乎没有运动。然而,在几毫秒内,meta I在热上放松到被称为meta II的形式,它实际上包含了三种不同(尽管在光谱上是等价的)形式之间的平衡。在第一步中,发生了质子易位,使元I的希夫碱去质子化,并使E113残基(109)质子化,生成元IIa。在第二步中,meta IIa经历了螺旋6向外倾斜,打开了一个裂缝与g蛋白相互作用,创造了meta IIb。在第三步中,一个质子被取到' ERY '基序的E134上,锁定螺旋6与螺旋3的关系。这种形式被称为“meta IIb H+”,是具有酶活性的R*形式,可以非常有效地激活G-蛋白。然而,它不能通过光逆转回到静止的11-独联体配置。
Piechnick等人(110)最近提出了一个视蛋白/视紫红质分子在激活过程中构型变化的示意图模型;参见他们的图4。在他们的模型中,蛋白质一方面可以以“紧致非活性”的结构存在,无论是在无配体(如游离视蛋白)的情况下,还是在结合后独联体视网膜(视紫红质),或者它可以以“开放的活性”结构存在,要么在视紫红质异构化到元II(如R*)时,要么在无配体时(如视紫红质*)。所有人的出入口反式或11独联体视网膜只能从/到打开的活性结构,这种通路似乎是通过在类视黄酮结合袋和脂膜之间打开的类视黄酮通道发生。一个单独的亲水通道,允许在席夫碱水解/形成过程中水的进入/退出,也被建议同时打开和关闭。该模型似乎能够解释许多已知的类视黄酮进入/退出、席夫碱形成/水解和蛋白质激活g蛋白能力的转换的特征。在第6节中,我们将指出视紫红质和视锥视蛋白之间的一个区别是水的接触更大,并且在视锥色素的“致密非活性”结构中仍然有可能。
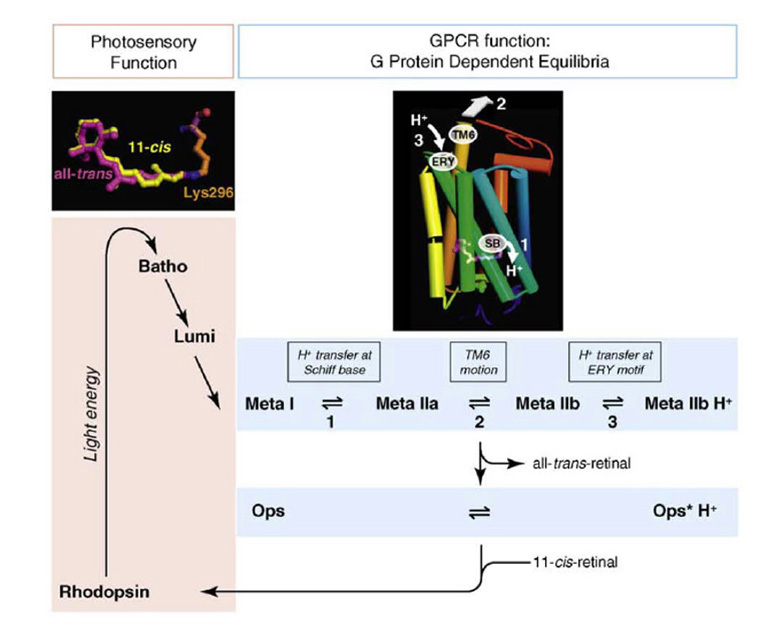
光活化后席夫碱键的快速水解。最近,Chen等人(104)表明,重新定位的反离子(相当于牛视紫红质的E113)的存在对光激活后视紫红质的II状态下席夫碱键的快速水解至关重要(在锥状态下发生水解的速度比棒状后视紫红质的II快约200倍)。如上所述,在后视紫红质的II中,席夫碱键是去质子化的,而反离子位点是质子化的。然而,希夫碱键的水解需要其瞬态质子化,Chen等(104)表明质子供体为质子化反离子残基;水就能攻击质子化的希夫碱,破坏共价键,从而留下所有的-反式视网膜非共价附着在类视网膜结合袋。因此,我们可以得出结论,反离子位置的重新定位有助于产生一种能迅速释放其全部-的视觉色素反式视网膜,因此有助于快速关闭锥体中快速反应所需的激活,也快速准备好视蛋白结合11-独联体视网膜。正如我们将在第6节中讨论的那样,在进化的后续阶段,Rh1视红质(视紫红质)找到了一种防止meta II快速衰减的方法,从而使激活的视紫红质集成光子触发信号的时间比快速衰减的meta II所能达到的时间更长。
c -视蛋白的脊椎动物前进化情景
根据以上信息,我们可以假设脊索动物谱系中c -视蛋白的进化过程中发生了以下情况(根据Terakita, Shichida和同事的解释进行了轻微修改):
C-1)一种古老的脊索动物具有一种c -视蛋白光色素,与现存的Opn3(脑视蛋白和tmt -视蛋白)、刺丝动物和原气孔c -视蛋白甚至r -视蛋白具有密切的同源性。E181是席夫碱反离子,113位点不带负电荷,可能是Y或f。这种光色素是双稳态的,可以从活性状态发生光逆转。它激活了一种Gi蛋白,尽管其激活Gt的速率比现代锥视蛋白或杆状视蛋白低得多。
C-2) c -视蛋白在113位获得谷氨酸。随后这个E113在暗静息状态下对希夫碱起反离子作用,而古老的E181可能在第一个后视紫红质中间体—后视紫红质I中起了稳定质子化希夫碱的作用。
C-3)进一步的突变发生,使得螺旋6在活性状态下更大的倾斜,从而改善了与g蛋白的相互作用。这种新的结构通过ERY基序的质子化来稳定,创造出高度活跃的后视紫红质II,这是脊椎动物“视觉”视蛋白所特有的。与前一种状态相比,这种后视紫红质状态表现出对g蛋白非常有效的激活,但在吸收另一个量子时不再能够发生光逆转(在紫外线中也有其吸收最大值)。
C-4)反离子位点的重新定位也加快了激活状态下席夫碱键的水解,提供了较短的meta II生命周期,因此在早期球果中有更快速的光响应。
C-5)脊椎动物视蛋白Opn3、顶视蛋白、副视蛋白、VA视蛋白,可能还有视蛋白,代表了在这一进化过程中出现的中间形式的现代版本,并表现出与祖先脊索动物视蛋白的功能属性不同程度的修饰,这些功能属性在不同类型的感光细胞中仍有应用价值。
C-6)因此,原始脊椎动物侧敏器官第一锥体光感受器的祖先视蛋白表现出g蛋白的高效激活和快速关闭;也有可能在这个阶段锥的g蛋白已经进化成Gt(转导蛋白)。
C-7)正如下一节将讨论的那样,这第一个锥视蛋白复制,产生了锥视蛋白的SWS和LWS(短波和长波敏感)分类。
C-8)这些事件都发生在脊椎动物辐射基地附近的“2R”全基因组复制之前。
6脊椎动物视蛋白“视觉”
颌骨脊椎动物视网膜的一个特征是使用锥视蛋白和锥视蛋白为基础的通道来接收从黄昏到黄昏的亮度(光视视觉),而视紫红质和杆状细胞为基础的通道来接收最低的亮度(暗视视觉),这在历史上被称为视网膜的“双重”组织。理解这种分裂是如何、何时以及为什么产生的,是很有意义的。与发生在茎类脊椎动物谱系中的许多其他因素一样,促成锥和杆的二元性的一个主要因素似乎是发生了两轮全基因组复制(“2R”),这是Ohno(111)最初提出的,随后在许多研究中得到了证实。
作为背景,我们将首先回顾锥视蛋白和杆视蛋白的系统发育。最近,通过对染色体上基因排列的研究,人们在理解“2R”复制的作用方面取得了重大进展。在这里,我们将检查与视蛋白相关的结果。然后我们将检查锥视蛋白和视紫红质在功能性质上的差异。
除了视蛋白在视锥细胞和视杆细胞中表达外,脊椎动物视网膜还表达许多其他视蛋白,包括黑视蛋白和VA视蛋白。此外,已有研究表明,表达黑视素的本敏光敏视网膜神经节细胞(ipRGCs)包含许多具有不同功能的亚型(112,113)。不同的亚型从不同的内丛状层亚层接收突触输入,并投射到不同的脑区。一些亚型能够促进小鼠的形态视觉。然而,下面关于脊椎动物视蛋白的讨论将仅限于视蛋白的视锥细胞和视杆细胞。
脊椎动物视蛋白的系统发育
发展史。视蛋白的系统发育,包括脊椎动物的“视觉”视蛋白,已经在许多出版物中进行了综述,包括Terakita (25), Bowmaker (114), Shichida & Matsuyama (8), Davies等人(94)和Porter等人(12)等。
锥视蛋白是视紫红质的祖先。关于脊椎动物视网膜视蛋白起源的最重要的发现之一是Okano等人(115)的发现,他们确定地表明杆状光色素视紫红质是从一种已经存在的锥状光色素进化而来的。此外,他们还表明,脊椎动物祖先视视蛋白的第一次分裂产生了SWS和LWS的区分;即一方面对长波敏感(LWS)锥视蛋白,另一方面对所有其他锥视蛋白和视紫红质(“SWS”部分,现在包括SWS1、SWS2、Rh2和Rh1)。这些基本的结果,由图22中最近的系统发育所说明,已被所有后续的研究所证实。
SWS/LWS分裂的时间。下一节将给出的证据(关于染色体的平行排列)表明,在脊椎动物谱系的基础上发生“2R”全基因组复制之前,祖先的SWS和LWS视蛋白就已经存在了。这两种锥视蛋白已经表现出了高效的转导激活,这是席夫碱反离子位置从181位重新定位到113位后的分子重排的结果。LWS红移灵敏度的出现似乎是这样发生的。
LWS视蛋白:通过氯化结合位点获得红移。由于残基E181不再需要作为静息态的反离子,突变不像以前那样受到限制。E181到H181的突变产生了一个氯离子结合位点,使得视蛋白的吸收峰发生了大幅度的红移,从而产生了LWS锥视蛋白。(注意,这个编号是相对于牛视紫红质的;在human OPN1LW框架中,残基为H197/K200)。Yamashita等人(116)最近的研究给出了对这种氯结合位点性质的研究历史。Wang等(117)研究表明,H181和K184两种残基对红移效应有贡献,其中H181为主要残基;K184可能有助于增强氯离子结合的稳定性。Davies等人(118)和Yamashita等人(116)最近表明,另外两个残基289和292也与氯效应有关。
另外,有趣的是,少量哺乳动物LWS锥视蛋白通过氯结合位点的丢失,随后恢复到蓝移的“MWS”敏感性。一个例子是小鼠' MWS '视蛋白,其181位突变为酪氨酸(Y181);它没有显示氯化效应,因为位置289和292也都突变为丝氨酸(Yamashita et al, 2013)。另一方面,豚鼠“MWS”视蛋白(也含有Y181)保留了部分氯离子效应,因为289和292位置都被丙氨酸占据。
慢波睡眠视蛋白重复。原始SWS色素复制2次,产生4种视蛋白,在光谱的蓝/绿区域具有最高灵敏度;下一节将提出证据,表明这种四倍复制是脊椎动物“2R”全基因组复制的一部分。
脊椎动物锥视蛋白。由于这些复制的结果,第一批脊椎动物(在那个阶段是无颚的)拥有五种不同的锥视蛋白,这些视蛋白被传统地分类为SWS1、SWS2、Rh2、Rh1和LWS(图22)。尽管视蛋白的某些种类在不同的谱系中消失了,但这五种视蛋白都被原始脊椎动物的所有后代继承了下来。例如,单孔目动物失去了SWS1视蛋白(119),而胎盘哺乳动物失去了SWS2和Rh2视蛋白;更多信息,参见Davies等人(120)。
在现存的脊椎动物物种中,视蛋白四类SWS1、SWS2、Rh2和LWS继续在视蛋白锥中表达。在随后的某个阶段(章节11),表达Rh1视蛋白的锥状光感受器进化为杆状光感受器。第二种杆状体存在于两栖动物中,利用SWS2视蛋白,但其起源的时间尚不清楚。
视蛋白基因的平行排列
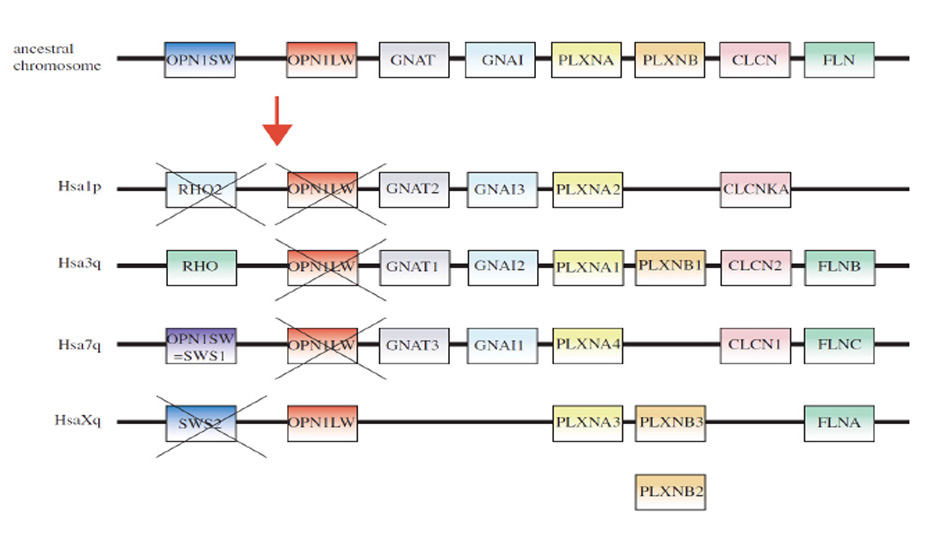
为了帮助确定编码视蛋白和光转导相关蛋白的基因的进化中全基因组和其他大规模复制的作用,Larhammar和同事研究了基因的平行子排列(7,121)。术语“平行染色体”适用于(通常)基因组中相关染色体区域的四重奏排列,在两轮基因组复制后约5亿年后仍然存在(尽管经常大大修改)。
视蛋白的染色体排列。Nordström等人(121)和Larhammar等人(7)对人类视蛋白基因(以及α亚基转导基因)的平行子排列的结果如图23所示。视紫红质(=RHO)、SWS1 (=OPN1SW)和LWS (=OPN1LW)的基因位于染色体3、7和x上的相应位置。鉴于脊椎动物视视蛋白的系统发育树总是以SWS和LWS视蛋白为最基础的分支(第6节),他们提出祖先基因(在四倍繁殖之前)的位置如最上面一行所示。在两次基因组复制之后,祖先SWS被认为进化为基础脊椎动物的SWS1、SWS2、Rh2和Rh1视蛋白,其中Rh2和SWS2在哺乳动物中已经丢失(第一列交叉)。此外,LWS视蛋白似乎只保存在四种视蛋白中的一种(现在人类的X染色体上)。
参与光转导级联的所有其他蛋白质的基因染色体排列将推迟到第8节。
观察到的系统发育树与“2R”重复预测的差异。重要的是要认识到观察到的脊椎动物锥/杆视蛋白系统发育树和全基因组复制树之间的差异。根据第一原理,人们会认为这对“2R”全基因组复制会产生一个具有双分支的“分叉”系统发生树,其形式为(SWS1,SWS2), (Rh2,Rh1),与通常观察到的“嵌套”树(SWS1,(SWS2,(Rh2,Rh1))相反。事实上,在四倍复制事件中幸存下来的所有基因家族都可能会出现双分叉树,而这种形式的普遍缺失最初被一些人作为反对“2R”复制发生的证据。
然而,如果(正如通常认为的那样)两个重复事件发生的时间相对于从那以后所经过的时间来说非常接近,那么各种各样的影响可能会改变所获得的系统发育树的形式。一般来说,任何在不同分支中加入不同替换率的倾向都可能导致提取的树拓扑结构的扭曲。对于视觉视蛋白,它们采用了不同的光谱位置和不同的动力学特性,在不同的分支中可能存在这些(或其他)特性之间的竞争,这在实践中导致了不同分支中残基的不同取代率。
举个例子,如果原始的SWS视蛋白在接近紫外线的地方显示其吸收峰值,并经历两次连续的复制,那么可能会发生什么。在第一次复制之后,似乎有可能是选择性压力导致了两种视蛋白的峰值波长分离。然而,这种压力很可能是不对称的,因为峰值灵敏度进一步向紫外线转移不太可能对生物体有利,而视蛋白中的一种转移到更长的波长(以占据SWS和LWS之间的间隙)则可能是有利的。类似的论点在第二次复制后适用。因此,可以设想,选择性压力最初会有利于全面转移分开不同视蛋白的光谱吸收峰,尽管在UV端有一个“屏障”。在此基础上,无论哪一种视蛋白最接近光谱的UV端,都将受到不改变其吸收峰的选择压力,因为任何方向的改变都将被证明是不利的。因此,与其他视蛋白相比,我们现在称为SWS1的视蛋白在影响其光谱调谐的残基上的取代率可能会更低。这可能会扭曲提取的系统发育树,使SWS1视蛋白看起来比实际更古老。
因此,观察到的视觉视蛋白的嵌套系统发育似乎不能作为否定这一假设的基础,即四种SWS视蛋白(SWS1、SWS2、Rh2和Rh1)起源于脊椎动物谱系基础附近的“2R”全基因组复制对。但另一方面,也没有证明' 2R '确实解释了五类脊椎动物视蛋白的存在。
锥视蛋白和杆视蛋白功能性质的差异
Imai等人(92)综述了锥视蛋白和视紫红质的功能特性差异,以及这些差异对氨基酸序列的依赖性。三个主要的区别包括:(我锥体后视视紫蛋白的寿命很短,(2)锥体色素的快速再生,以及(3)锥视蛋白对羟胺攻击的高敏感性。
锥元寿命短。许多研究表明,锥体视蛋白的meta II状态的寿命比视紫红质短几个数量级。Shichida的小组(在Imai等人(92)中进行了综述)在一定的温度和其他条件下,使用分光光度法检测了从天然膜或点突变培养的视蛋白中纯化提取的视蛋白的性质。对于WT鸡视蛋白,锥视蛋白的meta II衰变时间较短(SWS1、SWS2、Rh2和LWS的~2、~1、7和16 s),而视紫红质则长得多(210 s),每种情况下在室温下并在CHAPS中提取。最近,Chen等人(104)报告了类似的差异非洲爪蟾蜍其中蓝色/紫罗兰敏感(SWS1)锥视蛋白的meta II在20℃和pH 6条件下衰减,时间常数为~3 s,而牛视紫红质(Rh1)的时间常数为~800 s,提取液为0.1%十二烷基麦芽糖。他们的方法包括荧光显微镜测量来监测所有-的释放反式视网膜,推测反映元II的衰退。
观察到的meta II衰变(或视网膜释放)速度的巨大差异不是由于提取/纯化,因为Golobokova和Govardovskii(122)使用快速扫描二色显微分光光谱仪在完整的金鱼个体锥和杆中发现了类似的效果。他们发现红敏(LWS)和绿敏(Rh2)锥视蛋白的元II寿命为~5 s,而视紫红质(Rh1)的元II寿命为~330 s。因此,每一种方法都表明,视锥视蛋白的meta II衰变速度比视紫红质快100倍(或更多)。
地点122和189的作用。Yoshinori Shichida的团队已经使用位点定向突变来确定位点122和189(牛视紫红质编号)上的残基对meta II寿命差异以及色素再生时间差异的重要性(Imai et al .(92))。残基122在除深海鱼类外的所有视紫红质中被保守为E122,但在锥视蛋白中可能是I、L、M或q。残基189在所有锥视蛋白中被保守为P,但在大多数视紫红质中是I。这两个残基的突变在决定meta II的寿命和色素再生速度方面起协同作用。因此,棒状(E122和I189)的存在都有助于减缓meta II的衰减和视觉色素的再生,而锥状(I/L/M/Q122和P189)的存在都有助于加快动力学。总的来说,这两种残基的影响解释了meta II动力学中所有观察到的锥/杆差异。虽然有协同作用,但在两个位置上的效果并不一样强,在杆上122号位置的残留更有效,在球果上189号位置的残留更有效。
反离子位点的作用Chen等人(104)最近的研究调查了反离子位点在meta II衰变速度中的作用。如第5节所述,他们表明,全反式视网膜从SWS1视蛋白锥释放的速度比从视紫红质释放的速度快约200倍。在非洲爪蟾蜍SWS1视蛋白的反离子残留是天冬氨酸(D108),它的去除(通过突变为丙氨酸)导致全反式视网膜释放极慢;需要注意的是,这种视觉色素的希夫碱未加盐,因此对紫外线很敏感,所以它需要紫外线而不是蓝光来激活。将天冬氨酸重新引入另一个位置(残渣85),也接近席夫碱,恢复了紫的吸收,此外恢复了所有-的释放反式视网膜。他们还比较了锥视蛋白和视紫红质的视网膜释放,当反离子从两者中去除(在牛视紫红质中通过突变到谷氨酰胺);测量需要更高的温度和更长的监测时间,但值得注意的是,两种突变视蛋白的释放速度同样缓慢。
从这些实验和进一步的实验中,Chen等人(104)得出结论,锥体视蛋白和视紫红质中席夫碱键的水解速率是相似的,不同的是共价键被打破后视蛋白释放视网膜的速率。这可能是因为发色团与视网膜结合袋相互作用的差异,或者是发色团穿越视蛋白能力的差异(即其可达性)。如第5节所述,他们提出了一种Schiff碱水解的分子模型,在该模型中,反离子位点的质子化残基作为质子供体,使键瞬间质子化,从而使其容易受到水的攻击。
易受羟胺攻击。锥视蛋白容易被羟胺降解,而大多数视紫蛋白则不能。(这适用于胎盘哺乳动物的视紫红质,尽管一些非哺乳动物和单孔目视紫红质已被证明是缓慢降解的;(123, 124))。羟胺攻击希夫碱键,并且能够在静息视蛋白和激活视紫红质的后视紫红质状态下这样做。羟胺攻击希夫碱的方式被认为与meta II的正常衰变过程中发生的方式大致相同,即通过键的短暂质子化,因此锥/杆视蛋白对羟胺攻击敏感性的差异仅仅反映了结合位点对羟胺的可及性。
限制焦视紫红质II衰变和形成速度的分子机制.后视紫红质II状态的衰变涉及席夫碱键的水解,然后释放所有的反式视网膜从发色团口袋中分离出来,对于其中哪一个代表速率限制步骤一直存在争议。结合Chen等人(104)和Piechnick等人(110)的最新工作,提出以下几点:meta II态必须是瞬时质子化的,而质子供体是质子化的反离子残体。水是必需的,在锥视蛋白中相对不受阻碍,但在视紫红质中只能通过在meta II状态打开的亲水通道发生。同样地,当11独联体视网膜结合形成视紫红质,水分子必须能够通过同样的途径离开结合部位。在Piechnick等人(110)的分子模型中,视紫红质的亲水通道允许在“打开的活性”结构中缓慢地访问/去除水,但非常有效地保护了“紧凑的非活性”状态下的结合位点。对于锥视蛋白,有必要假设在活性和非活性构型中对水的获取限制都要小得多。
视紫红质难接近。因此,锥视蛋白和视紫红质之间的三个主要区别(meta II寿命、再生时间和羟胺敏感性)可能都只是表明,视紫红质分子在“屏蔽”视网膜共价结合位点上更有效,不受小分子(包括水和羟胺)的影响。此外,这种主要的锥杆差异似乎是由位点122和189的残基造成的。换句话说,这些残基的获取可能允许视紫红质分子采用更紧凑的“屏蔽”结构,在静置状态下不允许水访问,而在meta II状态下允许缓慢访问(见Piechnick等人(110))。因此,锥视蛋白和杆状视蛋白之间所有重要的分子差异似乎完全有可能源于可及性的差异,主要由这两个位点的残基决定。
锥视蛋白和视紫红质之间差异的演化。就进化而言,我们可以用以下观点(如第5节所述)对上述发现进行合理解释:视锥细胞的原始c视蛋白已经进化到其反离子位点的重新定位,这使得g蛋白激活的效力增加和全反式视网膜的释放,后者在一定程度上有助于快速反应。但随后(在' 2R '复制之后)增加敏感性的压力为其中一种视蛋白获得激活meta II的更长的寿命提供了优势,这在Rh1视蛋白中完成,当位点122突变为谷氨酸,位点189突变为异亮氨酸,它们在视紫红质的目前形式。
此外,缓蚀后视紫红质II可能还提供了一个优势,可以减少由all-的大量快速释放所引起的毒性反式强光(如阳光)中的视黄醛;如果是这样,这将为视紫质中E122和I189的保留提供压力。
深海鱼类中的残留122。有趣的是,在深海鱼类的视紫红质中,残留122经常被谷氨酰胺(Q122)占据,这在吸收光谱中提供了显著的蓝移(125)。可以想象的是,在极深的深度所经历的极低的光强度导致视网膜被释放的如此之少,因此在这些物种中E122Q从来没有毒性问题,而转移到峰值吸收的较短波长有一个优势。
相似点。锥体和杆状光颜料之间的一个有趣的相似之处,而不是不同之处,是它们在自发热异构化到活性态方面都非常稳定。这种稳定性的比较细节将在第10节中讨论。
脊椎动物视蛋白进化的情景
从目前所提供的信息来看,以下是导致脊椎动物视网膜中出现四视锥蛋白和视紫红质的进化事件的顺序:
D-1)在大规模复制事件发生在脊椎动物谱系的基础之前,中枢神经系统的吻端区域(会随着间脑的扩大)包含了表达一系列c视蛋白的神经元(其中一些是纤毛光感受器),包括视蛋白类的成员,视蛋白类的parietopsin,副视蛋白,VA视蛋白,视蛋白和祖先锥视蛋白。
D-2)在祖先锥视蛋白(和一些早期的c视蛋白)中,反离子位置已经迁移到113号位置。随后,发生了允许螺旋6更大运动的变化。新的meta II形式表现出了对g蛋白的高效激活作用,并能迅速衰变释放全反式视网膜。
D-3)这种古老的视锥视蛋白复制形成两个部分,SWS和LWS视蛋白,分别在两类独立的视锥光感受器中表达,位于横向扩展的间脑部分。
D-4)在LWS视蛋白中,原始的反离子位点E181(牛视紫红质编号)发生了突变到H181,形成了一个氯离子结合位点,从而使色素的吸收峰值发生了显著的红移。
D-5) SWS和LWS视蛋白的基因在一条染色体上相邻。不远处是Gt(转导素)和Gi的α亚单位基因。
D-6)经过两轮全基因组复制,产生了四个SWS基因的拷贝,这些拷贝经过突变,形成了SWS1、SWS2、Rh2和Rh1视蛋白。当绑定到11-时独联体视网膜,这些视觉色素的光谱灵敏度峰值从紫外到光谱的绿色区域。在复制产生的四个LWS基因中,只有一个存活了下来,对应于所有现存脊椎动物的LWS(有时也称为MWS)基因。
D-7)经光子激活后,所有锥视蛋白均表现出高效的g蛋白转导活性;当它们从元I态放松到元II态时,没有一个表现出光逆转;它们都经历了相对快速的热衰变,从活性元II状态释放所有-反式视网膜。
D-8)在Rh1视蛋白中,发生了两种突变(在残基122和189处),导致meta II中间体的衰减变慢。这使得表达Rh1视蛋白的光感受器能够更长时间地整合光子信号,从而赋予其更高的灵敏度;但代价是重新绑定11-的速度较慢独联体视网膜,因此视觉色素再生较慢。几乎没有其他东西可以将这种Rh1“视紫红质”与其他“视视蛋白”区分开来。
D-9)这些视蛋白抗异构化的热稳定性(即它们避免自发进入活性全反式态的能力)是它们被调谐到的吸收波长的函数(见后面的第10节),但所有这些视蛋白都是高度稳定的。
7脊椎动物视网膜视锥和视杆
纤毛细胞作为脊椎动物主要光感受器的胜利
鉴于大多数原气孔动物的成像眼都是利用微绒毛光感受器,我们很自然地会问,为什么脊椎动物的眼睛会选择纤毛光感受器。此外,鉴于单一类型的苍蝇横纹肌光感受器可以可靠地检测单个光子,并且在很大的强度范围内也能非常迅速地做出反应(Yau & Hardie(126)),我们也应该问,为什么脊椎动物使用具有独立锥和杆分裂的双视网膜。
在考虑纤毛和微绒毛光感受器的相对优点,以及这两类光感受器在不同生物体中占优势的可能原因时,重要的是要避免简单地比较生物体光感受器之间的特性的错误。例如,将现代苍蝇横纹肌光感受器的特性与现代锥状和杆状光感受器的特性进行比较是不恰当的。相反,我们需要考虑光感受器的特性,以及有机体在进化的相关阶段的环境,当细胞类型获得优势;也就是说,当光感受器的类别被“选择”的时候。在睫状体光感受器首次在脊索动物视网膜中占主导地位的阶段,超过500毫亚,证据表明杆状细胞还没有进化。同样地,在原始口复眼中,可能超过550 Mya的微villar光感受器获得主导地位的阶段,它们不太可能已经成为真正的横纹肌光感受器,也不太可能已经进化出高度专业化(可能是独特的)的光转导级联果蝇眼睛(127)。因此,我们需要研究比较简单的纤毛和微绒毛光感受器的相对优点,这些光感受器很可能在古代就存在。
考虑到这些要点,我们可以列出纤毛光感受器可能比微绒毛光感受器有一些优势,在它们之间做出“选择”的时候:
E-1)首先,响应的极性(对光的超偏光和对暗的去偏光)意味着纤毛光感受器在古代生物中非常适合于检测阴影。阴影引起的去极化可能触发动作电位或增加突触传递素的释放,而不需要额外的逻辑,这可能对具有简单神经系统的动物有利。
其次,两类光感受器之间的增益可能没有什么差别。最近,Ferrer等人(46)表明文昌鱼微绒毛光感受器的转导增益远低于现代横纹肌光感受器的转导增益。然而,脊索微绒毛光感受器的定量反应持续时间极短(几毫秒;(43))与纤毛光感受器较慢的反应相比可能是不利的。
E-3)第三,经计算,在明亮的环境中,离子电流在光中减少的超偏振光纤毛光感受器的能量消耗低于离子电流在光中增加的去偏振光微绒毛光感受器;另一方面,两类光感受器的能量消耗在黑暗中是相当的(128)。在明亮的环境中,双视网膜还有一个优势,因为视杆细胞饱和,从而将能量消耗降低到较低水平,只有视锥细胞继续产生循环电流,并随之产生能量需求(11)。
第四,脊索睫状体视蛋白(而不是其他睫状体视蛋白)发生了分子内的变化,大大提高了它们激活g蛋白的效力。结果,它们失去了进行光逆转的能力。
E-5)第五,这些新的纤毛视蛋白表现出相当迅速的活性后视紫红质II状态的衰减,释放出束缚的all-反式视网膜,从而使结合11-独联体视网膜。由于色素不能经历光逆转,这种释放是必不可少的;然而,也有可能这种变化先于激活功效的增加。在双稳态视蛋白(r -视蛋白和早期c -视蛋白)的情况下,被激活的色素在暴露于光后很长一段时间内保持后视紫红质状态,因此用来探测入射光子的视紫红质池被耗尽。在这方面,当生物从明亮的环境迁移到昏暗的环境并需要快速适应黑暗时,双稳态光色素可能是非常不利的。因此,脊索纤毛光感受器可能在这样的条件下提供了明显的优势。
E-6)最后,对生物体的额外好处可能不是直接来自于光转导过程的特性,而是来自于纤毛细胞进化出突触传递到它们的微绒毛对应体的事实(见第15节),从而通过单一的传入通路组成了一个双光接受系统。
无论如何,正如上面的讨论所显示的,有可能指出在古代脊索动物中纤毛光感受器可能优于微绒毛光感受器的几个重要方面。出于这样或那样的原因,纤毛光感受器确实在原始脊椎动物的感光器官中占优势。
多类脊椎动物视网膜感光器
关于视蛋白的分布、形态变化和跨脊椎动物光感受器转导途径的综述,见Ebrey & Koutalos(91)。
如第6节所述,这种古老的脊椎动物具有五种锥状/杆状视蛋白光色素。此外,它还具有锥体/杆状光感受器的五种形态纲。根据颌骨脊椎动物的光感受器的规范,以及现存七鳃鳗的间接证据(图15),似乎每一类光感受器都表达一种视蛋白。
在下颌脊椎动物中,这些光感受器包括四类视锥细胞,分别表达各自的视锥视蛋白(SWS1, SWS2, Rh2, LWS),以及一类单一的视视色素视紫红质(Rh1)。然而,至少在两栖动物中,SWS2“锥”视蛋白还可以在另一类视杆中表达(因其呈绿色而被称为“绿色视杆”)。在活的七鳃鳗中,所有五类光感受器都是锥状的,尽管表达“视紫红质”的那一类具有一些杆状的特性(第3节)。因此,“真正的”杆状细胞在图1第5点之前似乎没有进化出来。燃料棒出现的时间将在第11节中讨论。
目前尚不清楚锥体光感受器光谱类别的多样性的最初驱动力是否仅仅是为了覆盖更多的“可见”光谱,考虑到每个视蛋白吸收光谱的相对狭窄的区域,还是为了提供“颜色视觉”。但似乎有理由认为,几乎在生物获得新的光谱信息时,神经系统就会利用这些信息来提供颜色信息。
球果和杆之间的形态学差异
锥体和杆状光感受器的形态学细节在Webvision第一部分第2章“光感受器”中给出;这里我们将简单地集中在差异在两类细胞之间。
囊与椎间盘结构。哺乳动物视锥细胞和视杆细胞之间最显著的形态学差异与外节膜的拓扑有关。在视锥细胞中,整个膜与内段的质膜是连续的,而在视杆细胞中,外段的大部分膜以像泄气的气球一样的圆盘形式与质膜隔绝。这些盘实际上并不是“自由漂浮的”,而是被包括外周素/RDS、Rom1和环核苷酸门控通道GARP结构域在内的蛋白质系在质膜上。
在这两种细胞类型中,新的外段膜在纤毛颈附近不断形成,并向外凸起,因此新的囊在最近形成的囊下反复形成,Steinberg、Fisher和Anderson(129)的实验记录了这一机制;见上图16B。他们发现,边缘形成过程发生在相邻的囊膜之间,尽管在球果中,这一过程只在周围进行了一部分,因此囊总是对外开放的(专利)。然而,在棒状体中,这种边缘形成的过程沿着周长(双向地)进行,将相对的表面拉在一起,从而形成一个从质膜上夹下来的圆盘。
由于这种“封闭”的盘片在棒,新合成的蛋白质被困在新形成的盘片。随着更多的椎间盘形成,早期形成的椎间盘在被RPE吞噬之前,会沿着外节段的长度缓慢向外迁移,时间长达数周。蛋白质的这种定位和迁移可以通过应用标记的氨基酸(130)后连续时间的自放射检查来实验证明。相反,在视锥细胞中看到的是标签在整个外节段的均匀分布,如果蛋白质能够在外节段的连续质膜中扩散(甚至非常缓慢),这是意料之中的。
其他的形态学差异。它们的外部节段之间的进一步形态学差异是在杆上存在一个或多个切口;这些深入外节段的纵向凹痕增加了细胞质空间的横截面,增加了细胞内信使的有效纵向扩散系数(参见第11节)。在内段的水平上,锥体表现出棒状结构中没有的专门化,包括抛物面和有时的光谱过滤器(椭球体中的油滴或小泡)。可能是由于锥体的高折射率,锥体的内部部分非常有效地将入射光漏斗入外部部分,这导致锥体产生明显的Stiles-Crawford定向效应,而杆则产生较弱的Stiles-Crawford定向效应。在它们的突触末端,锥体和杆体有很大的不同,锥体显示大的椎弓根,而杆体显示小的球体。此外,球果和杆状植物之间还有其他微妙的形态差异。
视锥细胞和视杆细胞的功能差异概述
锥状光感受器和杆状光感受器彼此的光响应特性非常相似(见下文),细胞只表现出几个主要的区别:
(一个)棒.杆状光感受器响应的决定性特征是它的能力,在适应黑暗的条件下,对光的单个光子(131,132)的吸收做出可靠的响应(即具有良好的信噪比)。这种单光子探测性能是可能的,因为噪声,表示为“暗光”,在棒非常低;这种暗光在杆状体中比在LWS球果中低许多个数量级,尽管在SWS球果中差异可能更小。
(b)视锥细胞.锥体光感受器反应的决定性特征是它的能力:(我)迅速作出反应,(2)在巨大的强度范围内发挥作用,因此(3)在稳定的光线下,无论亮度有多高,它都不会饱和,而且(4)当强光熄灭时,它几乎能立即恢复大部分响应能力(在Lamb(133)中回顾)。
锥/杆相似之处。在大多数其他方面,锥状光感受器和杆状光感受器的反应非常相似,如表3所示,人类LWS锥状光感受器与杆状光感受器的比较。例如,在哺乳动物的视锥细胞和视杆细胞中,光转导级联扩增似乎相当相似。它们之间观察到的灵敏度差异(约20倍)在很大程度上是由于锥响应的更快关闭,在适应黑暗的条件下,锥响应通常比棒快5倍左右。如图24所示,锥杆响应开始上升时增益大致相似,但锥杆响应恢复得更快。除了时间轴的总体比例外,光响应的形状(即它们的动力学)也被证明是非常相似的。
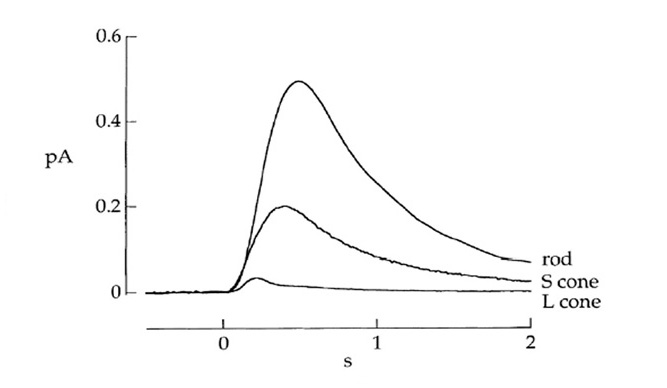
然而,视蛋白的转导增益和g蛋白被激活的视蛋白激活率在视锥细胞和视杆细胞中可能并不完全相同,确实有一些报告显示视锥细胞的增益较低。Kawamura & Tachibanaki(134)回顾了从鱼和蝾螈的孤立锥和杆状体中获得的测量结果,得出的结论是锥状体的增益要低得多。最近,Tachibanaki等(135)对光激活视觉色素g蛋白的激活率进行了细致的实验,发现在视锥细胞中g蛋白的激活率比视杆细胞低~5倍。然而,锥体生物化学测量中反复出现的困难是关闭反应如此之快,以致于在确定激活率时产生了问题。总的来说,锥细胞的转导增益可能是杆状细胞的0.2-1倍。
图24中的反应也说明了一般的观察结果,表达蓝/绿视蛋白敏感的视蛋白锥感光细胞(即具有SWS1、SWS2或Rh2视蛋白的视蛋白锥感光细胞)通常倾向于显示介于LWS视蛋白锥和视蛋白杆之间的响应特性;因此,蓝/绿敏感球果比红敏感球果更慢、更敏感。
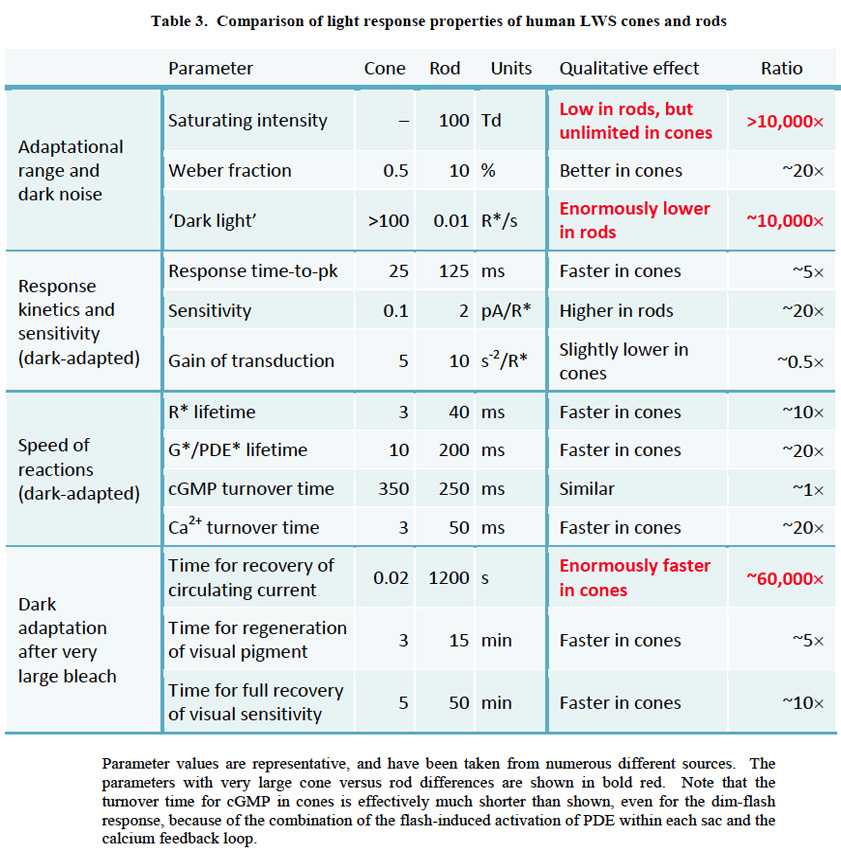
锥杆/杆的主要性能差异。在表3中,显示锥和杆之间主要差异的参数用红色表示。首先,尽管球果和杆状体都表现出经典的韦伯定律光适应,但对杆状体而言,这种适应只在有限的强度范围内发生,然后它们才会饱和,而对球果而言,这种适应持续到无限的高强度,因此它们在稳定的光中永远不会饱和。其次,光棒在黑暗中通常显示非常低的光子类事件的频率,即每几十或数百秒一个事件的顺序,而在LWS锥中,“暗光”可能通常是每秒数百个光子事件的顺序。虽然在黑暗光线中的这种差异很大程度上源于峰值吸收波长的差异(第10节),但一小部分似乎是由于视紫红质和锥视蛋白之间的内在差异。最后,在稳定的光消失后,使90%以上的光色素褪色,人体视锥细胞在约20毫秒(136)内恢复其循环电流,而人体视杆细胞的循环电流恢复可能需要20分钟,慢约60000倍。
这些差异将根据表现出优越性能的感光器类别来处理。因此,饱和的避免和响应速度将在第9节锥感光器中处理,而转导噪声和分解单个光子的能力将在第10节和第11节中处理。
8脊椎动物视网膜光转导级联的进化
在本节中,我们将研究脊索纤毛光感受器中使用的“脊椎动物式”光转导级联的进化,重点是脊椎动物视网膜的光感受器。在原气孔和刺鞘纤毛光感受器中使用的类似的级联只会在这里被提及。研究的目的是:(1)确定存在于我们脊索动物祖先身上的光转导级联的性质,就在全基因组复制的“2R”轮之前;(2)确定这种级联在锥细胞和杆细胞中专门形成的方式。因此,我们将研究级联的组成部分的共同进化,最初在脊椎动物之前的脊索动物中,然后在最早的脊椎动物中。
双侧光转导级联的演化
在图25A中,对比了几个光转导级联的主要成分,脊椎动物光感受器(上),其他纤毛光感受器和微绒毛光感受器(103)。每一个瀑布似乎都是从一个共同的祖先形态进化而来的。
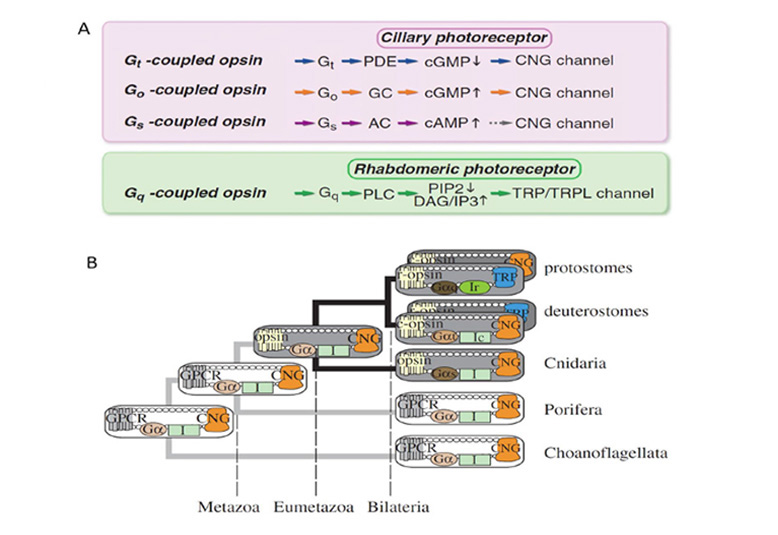
级联组分的协同进化。Arendt和Wittbrodt(137)研究了光转导级联过程中分子成分的共同进化,他们比较了视蛋白及其g蛋白、激酶和arrestin基因在原口动物和后口动物中的系统发育。他们发现了明确的证据,证明这四种成分肯定都存在于双侧动物的共同祖先身上。
耦合到g。视蛋白型与g蛋白型的偶联由Koyanagi等人(138)进行了研究,最近Terakita等人(103)也进行了研究。他们根据与视蛋白偶联的g蛋白将视蛋白分为四组。脊索c视蛋白与Gi/Gt偶联;至少有一个刺胞c视蛋白与Gs偶联;无脊椎动物“Go-opsins”夫妇去;而所有r -视蛋白都与Gq配对。
使用cngc的级联起源。Plachetzki等人(26,27)研究了视蛋白通过其g蛋白与离子通道的偶联演化。对于下游效应机制,在细胞的电响应水平上,Plachetzki等人(27)提供了祖先级联使用cngc(环核苷酸门控通道)的证据,如图25B所示。此外,替代TRP/TRPL(瞬时受体电位样)通道机制似乎出现在较晚的阶段,可能在双侧动物中,并在微绒毛光感受器中使用,其中r -视蛋白通过Gq耦合。
脊椎动物光转导级联的分子成分
(现代)脊椎动物光转导级联激活和恢复的分子和细胞机制在Webvision的“杆和锥的光转导”中有详细的处理,以及许多评论,包括Lamb & Pugh (139), Wensel (140), Yau & Hardie(126)等。所涉及的主要蛋白质,包括球果和杆状蛋白,如图26所示。关于这些蛋白质及其功能的概述,见Wensel (140);有关介导R*关闭的蛋白质的综述,见Gurevich等人(141)。
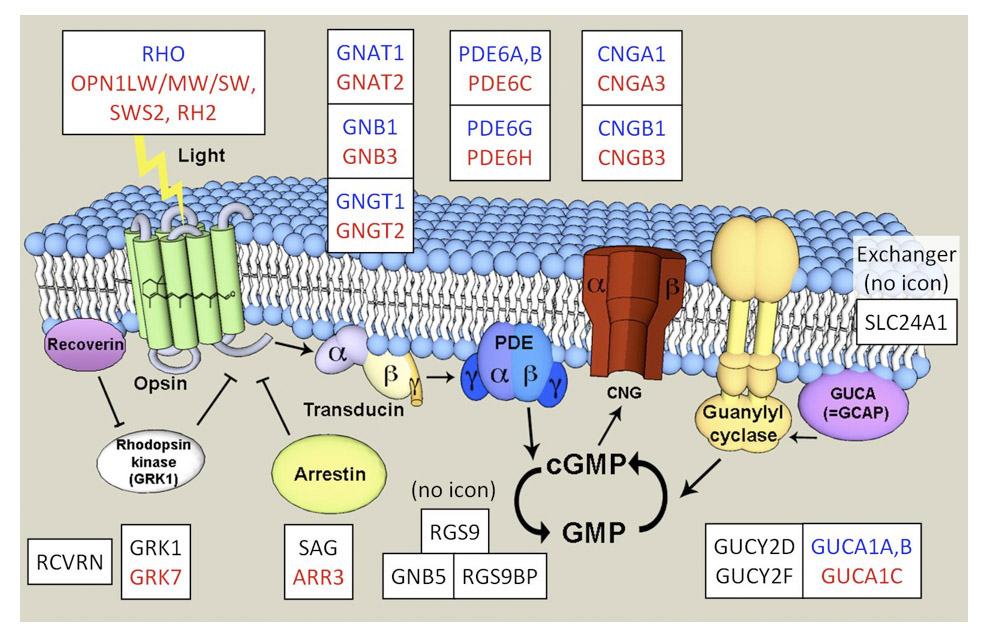
分子机制综述。光转导的分子机制在视锥细胞和视杆细胞中几乎完全相同。视觉色素分子的光异构化形成其活性形式R*,触发G蛋白(Gt,转导素)到G*的催化激活,进而激活cGMP磷酸二酯酶(PDE6)到E*。cGMP水解的增加降低了其细胞质浓度,导致环核苷酸门控通道(cngc)的关闭。这抑制了在黑暗中流动的循环电流,使电池超极化;此外,它还导致细胞质钙含量降低2 +通过Na的持续活动来集中注意力+/ Ca2 +K+交换器,Ca的下降2 +级别对于响应恢复很重要。响应关闭需要每一种激活形式的失活,以及cGMP水平的恢复。R*会被抑制素的结合迅速灭活,但这一步首先需要R*被g蛋白受体激酶(GRK)磷酸化。通过GTP酶加速RGS9-Gβ5-R9AP复合物的活性(GAP)作用,GTP结合到G*的末端磷酸被水解,G*/E*复合物迅速失活。钙的下降2 +浓度允许毫克2 +与禽酰基环化酶激活蛋白(GCAPs)结合,从而激活禽酰基环化酶(GC),恢复cGMP水平,从而导致离子通道重新打开。Ca的作用2 +赋予感光器一个强大的负反馈回路,帮助稳定电流。
锥状蛋白和杆状蛋白异构体和活性的差异
在视锥细胞和视杆细胞中调节光响应的蛋白质非常相似;事实上,两类光感受器中的一些蛋白质是相同的,尽管在大多数情况下表达的是不同的但密切相关的异构体,如图26和表4所示的基因名称所示。网上有许多用于检查与眼睛有关的基因的资源,其中有两个是有用的资源RetinaCentral.org(视网膜组或视网膜/RPE的转录组;(142))和RetNet, sph.uth.edu/retnet(视网膜疾病基因)。
当我们在第9节和第11节中比较锥和杆的功能特性时,我们将看到可以将杆转导级联视为锥转导级联的变体,其主要区别是在微弱闪光响应中的每个关闭步骤都被减慢了。有些光转导蛋白在视锥细胞中的表达水平远高于视杆细胞,这可能是光激活分子快速关闭的原因。一个显著的例子是分子复合物RGS9/Gβ5/R9AP在锥体中的10倍高表达水平,当它与PDE结合时,关闭激活的g蛋白转导。为了关闭激活的视紫红质,许多物种的视紫红质锥和视紫红质杆使用不同的GRK亚型(g蛋白受体激酶):视紫红质锥通常是GRK7,视紫红质杆通常是GRK1;在这种情况下,异构体的差异可能在很大程度上有助于减缓激活视紫红质的关闭,从而增加反应的敏感性(134,143,144)。对于微弱的闪光,恢复动力学也由cGMP在黑暗中的周转时间决定,而在棒中,周转时间减慢是由于棒PDE6的基础活性较低(145)。
Kawamura & Tachibanaki(134)和Korenbrot(144)对光转导蛋白不同亚型的活性和表达水平差异的总体影响进行了分析和建模,他们能够很好地解释观察到的球果和杆状细胞之间的动力学和敏感性差异。
脊椎动物光转导级联元件的演化
对于脊椎动物的光转导,Hisatomi和Tokunaga(146)比较了涉及的8个蛋白质家族的基因系统发育(转导蛋白,PDE, CNGC, GRK, arrestin, recoverin, GC, GCAP)。他们注意到基因树状图的分支模式非常相似,并得出结论,这些家族中的每一个似乎都进化出了“锥”和“杆”分支,尽管在两个案例(PDE和GCAP)中,杆状分支有进一步的重复。现在将介绍更多的最新结果,检查光转导级联的单个组件的进化。
Transducin。Muradov等人(87)研究了转导素α亚基的起源,他们在七鳃鳗中发现了两种异构体,分别命名为GαL和GαS,因为它们分别在长(锥形)和短(杆状)光感受器中表达。GαL与颌骨脊椎动物α亚单位的锥和杆转导的关系大致相同,可能代表了祖先的形式,而GαS进化支具有杆状版本,尽管它保留了某些锥样特征,如在n端附近存在“标志”的四残基序列。为了确定产生这两个转导亚单位的复制时间,还可以通过分析基因在染色体上的位置获得额外的信息(见下面第8节)。
磷酸二酯酶。用于脊椎动物光转导的磷酸二酯酶(PDE6)的独特之处就在于它能够受到γ亚基(Pγ)的调节,Muradov等人(86)和Zhang & Artemyev(147)研究了这两种成分的共同进化。Muradov等人(86)从七鳃鳗中克隆了这两种成分。对于PDE6催化单元,他们发现七鳃鳗有一个单一的亚型(如球果),与颌目脊椎动物的PDE6催化单元高度同源,尽管与球果和杆类动物的PDE6催化单元同样遥远,他们的结果与PDE6起源于脊索动物谱系中共同的PDE5/6/11祖先的概念一致。他们发现了一个与脊椎动物PDE6组合在一起的被膜PDE,但他们没有发现其他与PDE6组合在一起的非脊椎动物序列。
对于调节型Pγ亚基,他们发现了两种异构体,一种是锥状的,另一种介于锥和杆序列之间;在囊膜数据库中没有发现类似序列的迹象。他们的证据表明,这些调节亚单位出现在干脊椎动物谱系中,七鳃鳗和下颌脊椎动物的共同祖先很可能已经拥有两种亚型。Zhang和Artemyev(147)扩展了这一分析,他们提供的证据表明,尽管Pγ是严格意义上的脊椎动物发明,但PDE催化单元结合Pγ的能力早于抑制亚基的出现;事实上,他们的分析预测了刺丝动物的pde5 /6样酶应该与脊椎动物的Pγ相互作用。
中国兵器。在锥体中,环核苷酸门控通道由两个α亚基(CNGA3)和两个β亚基(CNGB3)组成,配置为A3-A3-B3-B3(148),而在杆状细胞中,该通道由三个α亚基(CNGA1)和一个β亚基(CNGB1)组成。Nordström等人(121)发现了有力的证据,表明产生环核苷酸门化通道α和β亚基的重复发生在原口动物和后口动物分化之前,他们还发现了暗示性的证据,表明α和β亚基的多个版本可能出现在“2R”重复中。
GRKs。Mushegian等人(149)最近研究了g蛋白受体激酶(GRKs)的起源和进化。他们发现有证据表明,GRKs起源于后生动物之前,通过激酶(类似于核糖体蛋白S6激酶)插入到与RGS (g蛋白信号调节器)同源的结构域的环路中。在脊索动物进化过程中,一个原始的GRKa分裂为GRK1/7和GRK4/5/6谱系。Mushegian等人(149)认为,这与第一轮“2R”全基因组复制相一致,尽管分裂可能发生得更早,因为被膜中存在一个明显的祖先GRK1/7Ciona intestinalis。Larhammar等人(7)的结果(见下一节)表明,GRK1和GRK7之间的区别是在“2R”重复过程中产生的。然而,还需要进一步的研究来解决这些异构体的起源问题。
Arrestin。Gurevich & Gurevich(150)和Alvarez(151)研究了抑制素(包括“视觉”抑制素)的系统发育,Gurevich等人(141)对这些蛋白质的功能进行了全面综述。抑制素在后生动物进化的早期就出现了,这个家族包括β-抑制素(得名于它们与β-肾上腺素能受体的相互作用)和“视觉”抑制素,在前双边动物时代从一个更大的家族中分化出来(151)。Kawano-Yamashita等人(152)为最近脊椎动物视觉抑制素和β-抑制素之间分裂的本质提供了线索,他们报告松果体非漂白视蛋白,副视蛋白,似乎通过内化过程被β-抑制素灭活(就像其他β-抑制素一样)。他们指出,Ciona intestinalis光感受器显然使用β-抑制素,他们提出脊椎动物式的视觉抑制素是专门用来关闭释放类视黄酮的视蛋白的;特别是脊椎动物视视蛋白。
光转导基因的染色体排列
视蛋白基因的平行子排列在第6节中介绍。在这里,我们考虑Larhammar等人(7)关于光转导的其他组件的平行子排列的结果。
Transducinα亚基。有趣的是,Larhammar等人(7)表明,α亚单位的转导基因与视蛋白位于同一邻位,而且非常接近(见图23)。最近,Lagman等人(153)将三聚体转导素的三个亚基的基因排列分析与基于序列的分析结合起来,他们提供了证据,证明所有三个转导素亚基家族在早期脊椎动物四倍化过程中都扩大了。他们的结论是,早期脊椎动物的四倍化为随后的转导亚基专业化提供了基础,导致锥体和杆状细胞之间的差异表达。
原始的GNAT基因可能是在双侧动物中存在的祖先GNAI/GNAT基因的后生口谱系中复制而产生的,因为一些原口动物有GNAI的同源物,但没有一个原口动物有GNAT的同源物;或者,复制可能发生得更早,但GNAT在原气孔中丢失了。
光转导级联的其他组件。Larhammar等人(7)还发现,在转导β亚基、GRKs和抑制素基因的局部组合中存在明显的平行子排列残余(图5)。此外,他们还发现了PDE6基因“2R”扩增的证据,CNGC通道的α亚基和β亚基以及抑制素和GRKs的证据。
解释。许多参与光转导级联的蛋白质,在视锥细胞和视杆细胞之间的异构体分布存在差异(部分或完全),Larhammar等人(7)研究的13种蛋白质成分中,至少有10种异构体的差异分布是脊椎动物谱系基础的“2R”复制的结果。因此,几乎毫无疑问,锥体和杆状转导级联之间的大部分区别可以追溯到两种全基因组复制导致的基因突变约束的减少。
前脊椎动物纤毛光感受器及其转导级联的假设性质
从现有的证据来看,要描绘出从祖先的双侧光感受器转变为脊椎动物光感受器的详细事件序列似乎还不可行。然而,在“原锥”光感受器及其光转导级联的进化过程中,在脊椎动物谱系的基础附近发生“2R”全基因组复制之前,有可能提出以下已经达到的状态:
F-1)“原视锥”光感受器的祖先视锥蛋白与现代视锥蛋白、SWS和LWS视锥蛋白大致相似,特别是它进化出了激活g蛋白的高效功效。
F-2)原始视锥视蛋白的复制产生了一对视网膜视锥视蛋白:一个带有标准E181残基的SWS视蛋白,和一个带有H181/K184组合的LWS视蛋白,后者提供了一个氯离子结合位点,产生显著的红移吸收。SWS和LWS视蛋白表达在独立但非常相似的视锥细胞中。这些视锥细胞的输出信号可能被用来提供二色视觉。
F-3)参与级联激活的主要附加蛋白已经进化成:(我) Gt的祖先转导,具有独特的α, β和γ亚单位;(2)原始的PDE6环GMP磷酸二酯酶,包括一对相同的催化亚基和一对相同的调节(γ)亚基;和(3)一种四聚体环核苷酸门控通道,由两类亚基组成,α和β。
F-4) Gαt可能是在脊索动物进化过程中由一个共同的祖先Gαi / Gαt演化而来。在脊索动物进化过程中,PDE6起源于一个共同的祖先PDE5/6/11。PDE调控亚基出现在脊索动物进化的晚期,在被囊动物分化之后(即图1中#4之后)。
F-5)除了锥视蛋白本身外,参与反应恢复和调节的主要蛋白已经进化为祖先的“锥”成分,由基因GRK7、ARR3、RGS9、GUCY2D/F、GUCA1C、SLC24A1和RCV1指定。
F-6)所有这些成分的存在表明,响应关闭的发生方式与现代脊椎动物光感受器的响应关闭方式基本相似。
F-7)特别是Ca所需要的成分的共存2 +反馈环(GUCA1C, GUCY2D/F, CNGA3/B3)提供了强有力的间接证据,表明负反馈环稳定了循环电流,并帮助光适应,已经存在。
F-8)转导级联的所有成分都在原锥体纤毛向外侧辐射的片层囊膜中表达。
F-9)因为释放所有-反式来自激活视蛋白(视蛋白是光感受器的外段)的视蛋白的类维生素a需要11-的来源独联体这可能是由胶质细胞(Müller)提供的。
细胞已经进化出一个简单的谷氨酸突触,允许突触传递。
9视锥细胞:具有特殊性能的感光细胞
视锥细胞表现出高灵敏度、高速度和高对比度灵敏度
高灵敏度。尽管经常有相反的说法,锥体光感受器是高度敏感的,并且转导的放大似乎与棒相当相似。事实上,当人们在夜间观察一颗昏暗的恒星时,正是视锥细胞(和光系统)发现了这颗恒星。例如,在观察昴宿星(七姐妹)时,尽管人们会通过暗点视觉意识到星团在中央凹旁区域是模糊的,但当人们固定住单个恒星时,是视锥细胞在探测它们。杆状系统因其较宽的空间总和而更适合于漫射星团,而锥状系统因其较高的空间分辨率而更适合于探测单个恒星。单个视锥细胞的灵敏度足以探测到非常昏暗的恒星,但对于非常昏暗的恒星,使用中央凹旁视杆视觉可能更好。
高速响应。锥形光感受器的响应速度远高于杆状光感受器,且响应速度随背景强度的增加而增加。在明亮的照明下,人类光视觉系统能够分辨频率约为100hz(154)的正波或方波闪烁,其beplay体育公司中亮/暗周期各为~ 5ms。因此,当照明熄灭大约5毫秒的时间时,适应光的视锥细胞似乎能够产生可分辨的信号。
对比敏感度高。人体光(锥)系统能够检测到非常小的分数变化的强度。因此,在适应光线的条件下,观察者可以检测到0.5%的对比度;相比之下,斑点(棒)系统的对比敏感度至少差一个数量级,为>5%。
因此,锥体系统是非常敏感,非常快,并具有良好的对比灵敏度。
锥避免饱和:无限的最高工作强度
与杆状光感受器相比,锥状光感受器在相当低的强度下就会饱和,而锥状光感受器在稳定的光照下都能很好地工作——无论稳定的背景有多强(155,156)。尽管在强光照开始时,视锥细胞可能短暂饱和,但它们会迅速恢复,并能够发出增加或减少的信号,即使光照太亮,使大部分光色素漂白。为了解释这一点,Lamb & Pugh(139)比较了人类视锥细胞和视杆细胞的关闭反应;他们表明,锥体避免饱和的能力可以用活化锥体光色素的关闭速度快20倍和活化锥体Gt/PDE复合体的关闭速度快20倍来解释。
在人类感光杆中体内,循环电流在~70 scotopic trolands (600 R* s)的稳定强度下减半-1),完全饱和发生在约1000个scotopic trolands(约104R * s-1).如果人体视锥细胞和视杆细胞的转导激活增益是相当的,那么两个非常短的视锥时间常数将使半饱和和完全饱和所需的强度提高约400×,达到~24万和~4 × 10的水平6R * s-1在锥。另外两个因素是在球果中转导的增益可能略低,哺乳动物球果的cGMP门控通道(与杆状细胞的通道相反)显示出对cGMP的增强亲和力2 +落(157);这些因素将进一步增加使锥饱和所需的R*速率。现在,最高的稳定的光异构化在人体锥体中所能达到的速率,通过暴露在稳定的光线下,对应于色素再生的最大速率,Mahroo & Lamb(158)的研究表明,该速率为0.75%/s,在一个含有4000万视蛋白分子的锥体中,对应约300,000 R* s-1.因此,稳定光异构化的最大可能速率低于上述计算出的使锥体饱和所需的异构化速率,这意味着锥体永远不能被稳定光饱和。
更直观的理解方法是,非常高的稳定强度导致锥视蛋白漂白到一个足够低的水平,以防止稳定光的响应饱和。因此,光视觉系统能够适应(即适应)巨大的强度范围,从beplay体育公司月光照射的条件到最明亮的阳光照射的场景。
锥循环电流恢复极快
当人眼暴露在足以漂白视杆90%光色素的稳定光下时,视杆循环电流在光消失后需20分钟左右才能完全恢复(159)。相比之下,一个稳定的光漂白90%的LWS/MWS锥光色素不会饱和锥响应(见上文),大约一半的循环电流保留。当光熄灭后,锥体循环电流在~20毫秒内完全恢复,比棒(136)快约60,000倍。
就像锥体避免饱和的情况一样,使这种循环电流的极度快速恢复的分子机制在于失活步骤的快速。Kenkre等(136)估计,在强光存在下,关闭反应的时间常数如下:对于R*, ~5 ms;G*/E*, ~13 ms;cGMP周转约4 ms。在cGMP动力学和通道活化的简单模型中使用这些参数,他们能够准确地拟合锥电流恢复时间过程的实验测量。在视杆细胞中,电流的极慢恢复不是由于g蛋白级联中失活/关闭步骤的缓慢,而是由于视觉色素再生的缓慢,加上未再生视蛋白激活级联的事实;关于这种缓慢复苏的完整解释,见兰姆和普(160)。
锥体所需的形态学特征
锥光感受器的操作要求对细胞的形态特征有一定的限制。
外段表面体积比高。为了实现快速反应动力学,所有的回收反应都必须是快速的,包括钙2 +-介导的鸟酰基环化酶活性加速,允许快速重新打开环核苷酸门控离子通道。这意味着细胞质钙平衡的时间常数2 +浓度(τCa)一定很短。在哺乳动物(包括人类)的视锥细胞中,τCa据估计约为3毫秒(161,162),而两栖动物的杆状体的值为400毫秒或更多。模拟结果表明,if τCa没有保持像其他关闭时间常数中最长的那样短,那么响应就变得强烈的双相甚至振荡。
直接的分析表明,τCa应该与Na+/ Ca2 +K+但与细胞质体积成正比(见Lagnado et al(163)的eqn(15))。因此,外部段的高表面体积比(允许在单位体积的细胞质中有大量的交换分子)将提供短的时间常数τCa对Ca2 +因此对视锥细胞是有利的。此外,对于视锥细胞,外节段的钙缓冲能力需要保持在较低水平。
开放囊的功能。上述对高表面体积比的要求无疑为球果中含有视蛋白的膜保持与细胞外介质的接触提供了强大的驱动力,从而使外段膜不像杆状细胞中那样密封。在LWS视锥细胞中,保留开放囊的另一个驱动力可能与氯结合位点H197(在牛视紫红质框架中=H181)有关,它在光谱灵敏度上产生了显著的红移。这个位置在第二个细胞外环中,为了在膜的这一侧提供高(且稳定的)氯离子浓度,可能有必要使这个区域保持在细胞外,而不是像在椎间盘中发生的那样转化为椎间盘内。
锥体光感受器中囊结构的存在似乎可能限制膜基反应和单个光异构化引起的细胞质反应,使其仅局限于单个囊的区域;根据考虑的响应特性,这可能是有利的,也可能是不利的。
突触释放约束。为了使锥体光感受器能够快速传输它们的渐变信号(即模拟信号),突触囊泡的释放速率必须非常高。这是必要的,因为突触后信号需要在神经递质囊泡定量释放所固有的噪声之上被检测到。为了能够检测(比如说)1%的囊泡释放变化,必须使囊泡释放的平均数量达到1/(0.01)的量级。2,或10000。为了在(比如说)100毫秒内实现这一目标,囊泡的释放速率需要在10万个囊泡左右-1这是非常高的。因此,锥体的突触末端需要一个非常大的区域作为活性释放区,以达到这种囊泡释放的速度。作为比较,在棒状细胞中,对单个光异构化的响应要比细胞的基线噪声大得多,因此足以检测(比如说)10%的变化量级;此外,杆的响应比锥体的响应慢得多。因此,一个小得多的囊泡释放速率(因此一个更小的突触)足以从神经棒提供一个足够可靠的突触后信号。
脊椎动物视锥细胞光转导的改进方案
以下场景是关于脊索纤毛光受体的光转导级联的改进,发生在脊椎动物视网膜的早期进化过程中:
G-1)当脊索外侧视网膜开始传递空间(即视觉)信息时(见第17节),对于快速反应、高灵敏度、适应不同强度和高对比度的能力有很大的进化压力。
G-2)这些压力导致了梯级中每个回收步骤的加速;即:激活视蛋白的关闭,g蛋白/PDE的关闭,钙的周转时间2 +浓度,钙2 +离子通道重新打开的反馈。
G-3)为了弥补由于反应加快而不可避免地导致的敏感性降低,将会持续施加压力,要求激活视蛋白增强g -蛋白的激活效果。
G-4)考虑到多种锥的存在,并不是所有的锥都需要同样的速度或同样的灵敏度。波长较短(因此热稳定性更强)的视蛋白系统可以降低速度,以达到更高的灵敏度。LWS系统具有较高的固有噪声,更适合实现最快的性能。
G-5)为了响应能够迅速传递反应的强烈压力,也为了能够在光线水平(低对比度)的微小变化中发出信号,突触末端发生了变化,导致突触递质的释放率非常高。通过这种方式,即使是锥体细胞内电压的短暂而微小的变化也会导致突触后递质浓度的变化,这种变化可以在泡状释放所固有的噪声之上被检测到。
G-6)当第一批(无颚)脊椎动物进化时,它们的外侧视网膜具有五种光谱级的视锥光感受器,表现出异常的响应特性,与现代脊椎动物的视锥大体相似。
10噪声:热异构化和其他噪声源
本节检查光感受器反应中噪声(波动)的来源,从视蛋白的固有特性开始,然后在转导级联内。
锥视蛋白和视紫红质的热异构化
在考虑视锥和视杆之间的差异时,一个主要的差异与噪声水平有关:LWS视锥比视杆噪声大许多个数量级(表3)。在这里,我们将研究这种噪声差异在多大程度上是具有不同峰值波长的视蛋白分子的稳定性的函数。
60多年前,Stiles(164)开发了一个描述视觉色素光谱灵敏度长波长下降的模型,后来被Lewis(165)扩展,引用了分子振动的热能可以加入光子的能量以触发激活的概念。Barlow(166)扩展了这一概念,提出即使在没有光的情况下,自发热激活也可以以非常低的速率发生,并且他预测波长越长色素的稳定性(噪声越大)会降低。Barlow的预测已经在大量的实验研究中得到证实(167,168),并且已经提出了不同复杂性的理论模型来解释峰值波长和热激活速率之间的确切关系(165,168-170)。
一个重要的观察结果是,锥视蛋白可以非常稳定,当这种特殊视蛋白在杆状细胞中表达时,这一点最为明显。在两栖动物中,所谓的“绿杆”(吸收蓝色,因此呈现绿色)表达一种蓝敏感色素,它被证明是SWS2视蛋白,在同一视网膜的蓝敏感视锥中也表达(171)。早期的电生理学实验表明,蟾蜍的绿棒具有非常低的自发热异构化率(172),而最近的实验表明,这一率低得惊人,小于10-14年年代-1在23°C(168)。这意味着,当SWS2(锥)色素在蟾蜍杆中自然表达时,平均每10次才会自发激活一次14S,约4000年。因此,这种“锥”色素是所有已知视蛋白中最稳定的,包括传统的视紫红质!
锥:暗噪声
海龟LWS锥细胞内电压记录显示噪声水平非常高(173)。通常情况下,这种噪声的振幅会因“平均掉”而降低,这种“平均掉”是由于信号在电耦合网络中广泛横向传播的结果,而电耦合网络是由连接相邻锥(174)的缝隙连接所创建的。但在不与相邻粒子耦合的锥体(“电孤立”锥体)中,观察到的约3000 μ V峰对峰的暗噪声振幅与仅有25 μ V的平均单光子响应振幅相比是巨大的,并完全排除了单个光子事件的分辨率。计算表明,这种暗噪声相当于2000多个光异构化的真实光所产生的量子涨落-1;这个参数被称为锥体所经历的“暗光”。有背景的实验表明,应用这种强度的真实光,锥的闪光灵敏度大约减半。随后,在猕猴LWS/MWS锥细胞内测量中,Schneeweis和Schnapf(175)发现了他们计算的相当于>3000 R* s的暗噪声-1/锥。
LWS锥体细胞内电压暗波动的来源尚未确定。在海龟视锥细胞中,视蛋白的热激活有望产生显著的作用(见上文),因为吸收峰波长异常长,约为620纳米;事实上,这似乎是所有视蛋白中已知的最长波长的灵敏度峰值。如果所有的暗噪声都来自热异构化,则锥的视蛋白含量为~108分子除以2000秒的事件速率-1会产生平均热寿命0.5 × 105s,与Luo等人(168)的预测相差不远;图4C)表示该波长的锥视蛋白。除了光色素噪声外,转导过程也可能产生噪声(见下文)。最后,突触末端离子通道的抖振活动应该会产生额外的噪声。迄今为止,尚未对这些潜在来源的相对贡献进行评估。研究其贡献的一种方法是使用吸管实验,尽管一个潜在的缺点是锥体外段与正常的RPE细胞之间的纠缠分离可能会改变锥体的性能,与完整的视网膜相比。
棒:转导噪声和单光子响应的可变性
转导杆中的噪声。在视紫红质棒中,可以将转导过程中的噪声(以及随后的噪声)与视紫红质热异构化引起的噪声分离开来,因为自发的类光子事件是单独可分辨的(至少在适应黑暗的条件下是这样)。在适应黑暗的蟾蜍杆中,Baylor等人(176)报告了剩余的“连续”噪声对应于振幅约为单光子事件的1/400的射击事件,并以大约6000个事件的平均速率随机发生-1.从功能上来说,这个噪声源可以被认为是低的,因为尽管它存在,绝大多数的单光子响应都是可解析的。似乎杆状转导级联蛋白比锥状级联蛋白进化出了更强的抗自发激活的稳定性,Muradov等人(145)对PDE6的研究结果表明,杆状结构具有较低的基础水解活性,同时对转导蛋白的亲和力较弱(参见第8节)。
棒单光子响应的变异性。尽管在大多数关于该主题的出版物中描绘了相反的观点,棒的单光子响应是相当可变的,特别是就其时间过程而言,但大多数作者将动力学描述为“可复制的”。对于蟾蜍杆,Whitlock & Lamb(177)通过匹配以相似斜率开始上升的单个响应,直观地说明了单光子响应时间过程中的广泛变异性,并展示了峰值和下降阶段可能有多么不同。当他们估计一个单关步的变化时,需要解释实验测量,他们发现变化系数(标准偏差除以平均值)为~0.4。这是足够高的,它是不合理的描述定量反应显示“可复制的”动力学。在时间过程和振幅上的巨大变化是杆光响应中已知关闭步骤的完全正常(和预期的)特征,并已被Hamer等人(178,179)和Gross等人(180)精确模拟。
棒单光子响应的大小。最近的研究表明,在视网膜切片制备中的小鼠杆显示出比先前使用吸吸管方法报道的更大的单光子响应,在细胞中平均振幅为2-3 mV,最大响应约为20 mV(181)。有了这么大的单光子响应,可靠的突触传递响应到棒状双极细胞远没有文献中经常提出的问题。
11棒的特殊化和棒的起源
正如在第7节中所讨论的,杆状光感受器与锥状光感受器的决定性特征是其在黑暗适应条件下对单个光子(131,132)的吸收做出可靠响应(即具有良好的信噪比)的能力。尽管在形态学、分子成分和响应特性方面,锥状细胞和杆状细胞之间有许多不同之处,但从来没有一个令人信服的解释是什么使脊椎动物杆状细胞能够实现它们的光子分辨性能。
本节首先分析有助于棒可靠地信号单个光异构化的结构特征。然后研究了两种不同寻常的“杆状细胞”:一种是夜行的壁虎,它们的光感受器表现出杆状的特性,但相对较晚才从锥细胞进化而来;还有在鳐鱼的全杆状视网膜上,杆状细胞表现出某种锥状的特性。然后讨论了早期脊椎动物中杆状体的出现,最后描绘了杆状光感受器的进化场景。
结构作为一个因素,在棒的能力信号个别光异构化
为了达到一个合理振幅的单光子响应振幅(例如5%的循环暗电流),离子通道必须在中等长度的外段受到影响,这就需要细胞质信使cGMP沿外段纵向扩散。在典型的锥体中,在锥体响应的短时间内,细胞质信息主要被限制在单个囊的内部,不能提供超过1/N循环电流的单光子信号,其中N为外段囊的数量(通常为>500)。
更有效的纵向扩散(更高的纵向扩散系数)可以通过增加纵向扩散可用的截面积来实现,正如Lamb等人(182)最初所建模的那样。在锥体外段中,只有一小部分囊周长可作为细胞内的纵向通信通道,因此有效的纵向扩散系数很低。但是,通过将囊膜包裹在质膜内,通过形成夹断的椎间盘,基本上整个周长就可以作为一个纵向导管。纵向扩散系数可以通过使用一个或多个切口(外膜段的纵向内嵌)进一步增加,因为这将增加围绕椎间盘的间隙的总长度,因此将增加沿外膜段的细胞质路径的截面积。
Lamb等人(182)在蟾蜍的大棒中测量了激活的纵向扩散,在单光子响应峰值时具有3 μ m量级的长度常数,在光适应过程中测量了脱敏的稳态扩散,其长度常数约为6 μ m。随后的研究证实并将这些观察结果扩展到哺乳动物的杆状细胞。
有了胞质信使在外段纵向扩散的这种程度,该棒很容易实现5%循环暗电流的单光子响应振幅(前提是它的关闭反应已经足够慢)。Lamb & Pugh(70)模拟了这种纵向扩散对单光子响应时间过程的影响;附录B)。
夜行壁虎:具有杆状特性的球果
夜间活动的壁虎的光感受器提供了一个重要的测试,用来测试什么是制造“杆”的必要条件,因为本质上它们是带有一些杆状特征的视锥细胞,但它们表现出杆状的响应特性。Walls(183,184)提出,夜行壁虎的光感受器已经从球果“转变”为杆状。他认为现存的壁虎是由一种白天活动的祖先进化而来的,这种祖先拥有全锥体视网膜(即完全失去视杆的视网膜)。他认为,夜间活动的壁虎通过进化出杆状的外节来适应环境,同时保留了锥的其他形态特征和全锥视网膜的组织。在最近的文献中,大多数作者都提到了夜行壁虎的视网膜。Gekko盖柯)是“全杆状体”,感光体是“杆状体”,但这是不案例(见下文)。
光反应。光感受器在细胞内记录光的反应Gekko盖柯Kleinschmidt & Dowling(185),他们报告的响应特性与其他物种的杆状体中发现的响应特性大致相似。在25°C下,对微弱闪光的响应达到峰值的时间较慢,约为700 ms。尽管这些反应看起来非常敏感(因此像棒状),但我还无法提取出它们敏感性的定量值。对强度增加的闪光的反应族让人想起杆状族。对明亮闪光的响应振幅为~20 mV,从峰值迅速下降到平稳;用100 mM天冬氨酸处理后,这种凹陷被消除了,这表明它是通过水平细胞的反馈产生的;如果是这样,这方面将是锥形而不是杆状,因为杆状通常显示快速凹陷起源于内部部分。在稳定的背景下,光感受器的响应光适应,在~4 log的范围内10单位,在饱和;这种饱和度是杆状的。
Rispoli等人(186)报道了从孤立的外部节段的电压钳电记录Gekko盖柯光感受器,在连接纤毛处被分离然后用富含能量的细胞内溶液透析。平均暗电流为~67 pA,对增加强度的闪光的响应家族与其他物种的杆状体非常相似。在17°C下,对微弱闪光的响应达到峰值的时间为~1.1 s,报告了棒的动力学形式。通过计算暗适应闪光的灵敏度(根据强度测量),对应的是~0.8 pA的单光子响应振幅,而波动分析(集合方差除以平均值)给出的值略小,为~0.6 pA。这表明单光子响应相当于暗电流的约1%,远远大于传统的锥体,但略小于大多数杆体。
综上所述,这两项研究的结果表明Gekko盖柯光感受器对光照表现出主要的杆状特性,尽管单光子响应的振幅可能不像其他物种的真杆状那样大。
光感受器的形态。在一项超微结构研究中,Röll(187)确凿地表明,夜行壁虎的光感受器的几乎每一种形态特征实际上都是锥形的,证实了Walls最初的观点(183,184),并与最近关于它们是杆状的断言相抵触。唯一的杆状特征与外节段有关,并且:(1)外节段的大尺寸,(2)外节段“圆盘”的存在,大部分被质膜包围,(3)切口的存在。她在所有被研究的壁虎物种(包括夜行和昼行壁虎)的光感受器中显示出的锥状特征如下:连接的纤毛很短;内段包含一个富含糖原的抛物面和一个椭球(日生物种中有油滴);核内染色质分散;突触末梢与锥体蒂非常相似。她的结论是:“夜行壁虎的视网膜一定要归为锥状视网膜”。
夜行壁虎的光感受器外段呈盘状结构,此前有Yoshida(188)的报道,Röll(187)的研究结果非常一致。Yoshida(188)指出,“与其他脊椎动物的视棒一样,成年[夜行]壁虎视网膜中感光细胞外段的双膜盘堆栈被质膜包围,除了外段的近端区”。两项研究都发现绝大多数椎间盘被质膜包围,但也都报告了频繁的圆锥状开口,沿外节段的长度分布。两者都报告了一个(或很少有两个)棒状切口的存在。
Gekko视蛋白。锥视蛋白和杆视蛋白之间的区别在第6节中介绍。的两个主要观点Gekko盖柯已被证明与其他脊椎动物的LWS和Rh2锥视蛋白演化,并表现出锥状生化特性(189)。特别是,壁虎Rh2视蛋白具有Q122和P189,据报道,这两种视蛋白是锥体视觉色素快速元II衰变和快速色素再生特性的基础(92),第6节)。
壁虎phototransduction级联。转导的蛋白质级联进入Gekko盖柯Zhang等人(190)进行了研究,在他们能够识别的所有蛋白质中,序列是锥形的而不是杆状的;这些确定的蛋白包括Gt的α亚基(GNAT2), PDE6的催化和抑制亚基(PDE6C和PDE6H),环核苷酸门控通道(CNGA3)的α亚基和ARR3。虽然这些蛋白质主要是锥形的,但也有一些残基被鉴定为棒状的。另一方面,在测量表达水平和活性时,他们发现RGS9的表达水平低于杆状细胞的预期水平,而不是锥体细胞的高水平特征,由此产生的GAP活性也相应较低;同样,PDE的低暗基础活性是杆状细胞而非球果细胞的典型特征。
综上所述,视蛋白和级联蛋白的这些结果表明Gekko盖柯光感受器绝大多数是锥状的,但至少两种在产生慢反应中很重要的蛋白质(GAP活性和基础PDE活性)的表达水平和/或活性相反是杆状的。
滑板:能表现出锥状特性的杆
一些深水种的鳐鱼已经被证明拥有纯粹的视杆视网膜,但它们的视杆能够以锥状方式工作,其强度通常会使视杆饱和(191)。这些物种的棒子外段具有经典棒子的所有形态和其他特征,包括大的圆柱形外段、封闭的圆盘、500 nm处的显微分光光度峰和标记氨基酸的带状结合。另一方面,突触末端的超微结构介于常规杆和锥之间,有多个突触带,在内陷和平坦触点处都有多个突触触点(192)。
Dowling & Ripps(193)利用从离体视网膜中提取的天冬氨酸分离的跨视网膜电位记录了这些细胞的电反应,该电位反映了光感受器的活性。在暗适应条件下,电响应与常规棒非常相似。此外,应用稳定的背景照明导致饱和度在相对较低的强度,如在其他棒。然而,当这些背景持续一段时间后,神经棒慢慢恢复,并能够对增加的刺激做出反应。对于中等强度的背景,在5-10分钟内达到适应的稳定状态,但对于更强的背景,棒需要20分钟或更长时间才能从饱和恢复到稳定的适应状态。这些经视网膜记录的发现随后在适应黑暗条件和单一中等亮度背景强度下由滑板杆的吸液吸管记录证实(194)。吸液吸管的测量结果表明,当电池从初始饱和状态恢复时,闪现反应的加速涉及到每一个关闭反应的加速,这与暴露在稳定背景或温度变化下的传统棒发生的方式大致相同。
这些结果与滑板全杆视网膜中的杆在完全黑暗适应条件和昏暗照明下的功能与传统杆一致。然而,当暴露在更明亮稳定的背景下时,滑冰杆最初饱和,但在几十分钟后恢复,并显示出与锥细胞相似的适应特性。因此,在这种缺乏视锥细胞的视网膜中,视杆细胞似乎经过了某种程度的修饰,使它们的反应特性从杆状慢慢改变为更像锥状。目前,这种转变的细胞和分子机制尚不清楚。
在传统的“双”视网膜中,包括视杆和视锥,视杆在中高背景强度下不需要发挥作用。事实上,视杆细胞在视锥细胞工作的强度下达到饱和是有明显优势的,因为这减少了视网膜外部的代谢负荷,并允许氧张力维持在视锥细胞的合理水平。但在全杆状的鳐鱼视网膜上,杆状的视网膜似乎不得不放弃这种节能的技巧,以使动物能够继续看到在其他情况下会致盲的强度。
制作“真正的棒子”需要什么?
根据第10节和第11节给出的结果,对于制造“真棒”所需的关键特征,我们做出了以下解释。真棒被定义为能够可靠地发出单个光异构化发生的信号的棒,与具有锥状特性的光感受器不同:
H-1)首先,光感受器需要表达一个峰值光谱灵敏度小于约520 nm的视蛋白,以达到足够低的热异构化率;因此,LWS视蛋白被有效地排除在外。
H-2)光转导级联的放大不需要与传统锥体中的放大有显著差异。
一个基本的变化是,所有作为暗闪光响应恢复基础的关闭反应都需要足够慢,以便对单个光异构化的响应能够建立足够的抑制至少1%(最好更多)的循环暗电流。
H-4)与典型的锥管关断相比,所需的减速需要进行协调一致的改变:(1)减缓R*的关断;(2)减缓G*/E*的关断;(3)在黑暗条件下降低cGMP翻转时间;(4)减慢Ca2 +-介导的负反馈循环,从而减缓cGMP水平的恢复和离子通道的重新打开。这些变化是通过蛋白质活性的改变实现的,还是通过表达水平的改变实现的,这并不重要。
H-5)这种减缓的实际实现方式似乎包括:减缓GRK和抑制素磷酸化并与R*结合的速率;通过降低RGS9复合物的浓度来减缓GAP活性;通过降低PDE6的基础活性来减缓黑暗条件下cGMP的周转时间。
其次是细胞质信使蛋白(cGMP和Ca2 +)需要能够沿外段向轴向扩散大量距离,这需要外段形态的显著变化:囊膜需要被质膜包围(实际上形成圆盘),以一种方式为轴向运动提供一个扩大的导管。此外,外节段的一个或多个纵向切口可以进一步帮助轴向扩散。
H-7)一个额外的改变将有助于渲染(现在)在背景噪声之上可检测到的大单光子响应,这是通过增加级联蛋白的稳定性来降低光转导过程中的固有波动。
H-8)通过以上所有变化的组合,应该有可能将专门用于高性能快速光位视觉的锥型光感受器转换为能在黑暗适应条件下可靠地向单个光异构化发出信号的杆型光感受器。然而,关闭反应减缓的一个不可避免的后果是,这个细胞在低到中等背景强度下饱和。
棒子出现的时间
在第10节中,展示了现代SWS2 '锥'视蛋白具有极低的热异构化率,使它们特别适合作为两栖动物绿色杆的光色素。我们不能确定祖先的SWS2视蛋白是否同样稳定。但是,即使它的热稳定性差了好几个数量级,如果视蛋白在“棒状”中表达,那也完全足以支持可靠的单光子检测。我们可以得出结论,锥视蛋白(至少SWS2变种)在视紫红质进化之前就已经表现出足够的热稳定性,使单光子探测成为可能,因此可能在视紫红质杆进化之前。因此,视蛋白分子的热稳定性似乎不是杆状细胞进化的限制因素(至少,当“杆状细胞”具有探测单个光子的能力时是这样)。
很明显,在圆口动物从颌骨脊椎动物谱系分化之前,具有某些杆状特征的光感受器就已经进化了。因此,盲鳗光感受器据报道具有被质膜包围的盘,其光谱灵敏度峰值在500 nm处(章节3)。同样,一类七鳃鳗光感受器具有许多杆状特征,包括一种视蛋白的表达,该视蛋白似乎与Rh1同源,尽管该细胞在解剖学和电生理学上也保留了其他锥状特征(章节3)。这些盲鳗和七鳃鳗的光感受器是否能够对单个光子的吸收做出可靠的反应——换句话说,它们是否达到了颌骨脊椎动物“真棒”的决定性特征——的最高灵敏度。如果未来的实验能证明盲鳗和七鳃鳗的感光器都符合这一标准,那么这将成为支持这一观点的证据,即它们的共同祖先也拥有真棒。
在这个问题得到解决之前,我们不可能有把握地进一步说,首先,鳗和七鳃鳗的最后一个共同祖先似乎极有可能拥有一类具有许多杆状特征的光感受器;其次,“真杆状”肯定是在第一批有颌的脊椎动物出现时进化出来的。
“真杆状光感受器”出现的情景
以下步骤被认为是杆状光感受器从锥状光感受器前身演化而来的可能方法:
I-1)在“2R”轮全基因组复制之后,早期脊椎动物的外侧视网膜拥有五类锥状光感受器细胞,每一类都表达五类视蛋白中的一种(LWS, SWS1, SWS2, Rh2, Rh1)。
I-2)在表达Rh1视蛋白的光感受器中,逐渐发生了一些变化,导致反应变慢,同时增加了其灵敏度。
I-3)表达Rh1的光感受器的这些变化包括三个关闭反应的减慢和负反馈循环的减慢。其中一些变化涉及到该细胞中表达的关闭蛋白的特化,一些涉及到表达水平的改变,还有一些涉及到形态的改变。
I-4) R*关闭变慢,部分原因是残基122和189突变导致后视紫红质II的衰减变慢,部分原因是激酶和抑制素(GRK1和SAG)的变化。
I-5) G*/E*关闭变慢主要是由于RGS复合物(RGS9-Gβ5-R9AP)水平降低。
I-6)由于静息磷酸二酯酶活性降低(PDE6A, B),黑暗条件下cGMP的周转时间延长。
I-7)细胞质钙平衡的时间常数2 +通过钠的数量减少,浓度平行增加+/ Ca2 +K+部分是由于表面体积比降低,部分是由于细胞质钙的增加2 +缓冲能力。
I-8)外节片片边缘的变化导致质膜包围片片组(因此成为盘组),为细胞质中的轴向扩散提供了增强的截面积。
I-9)随着这些变化的逐渐发生,但同时发生,随着时间的推移,表达Rh1视蛋白的光感受器会变得越来越慢,越来越敏感。
I-10)最终,几乎整个外层部分被密封,提供了广泛的纵向扩展,关闭反应变得足够慢,以至于对单个光异构化的响应变得足够大,足以超过电池中的噪声水平。
关闭反应的任何进一步放缓都将是不利的。
I-12)这种细胞的突触具有要求较低的作用,因为信噪比较高,动力学较慢,因此突触能够在较少的突触机制消耗(即较少的带状和较少的囊泡)下令人满意地发挥作用。
I-13)这些综合渐进变化的最终结果是“真棒”光感受器的出现,它可靠地发出单个光异构化的信号。
12类维生素a在黑暗中的再异构化
正如第5节所讨论的,脊椎动物视网膜色素和无脊椎动物视网膜色素的一个主要区别是脊椎动物视蛋白释放它们的all-反式光激活后的类维生素a。因此脊椎动物的视网膜需要持续供应11-独联体为了抑制游离视蛋白的残余活性,同时再生原生视紫红质,以便通过光进一步传递信号。这是通过两个在黑暗中运行的生化途径完成的(图27;Lamb & Pugh, (160);Wang & Kefalov, (195);群(196))。一种是经典的“视黄素循环”(图27B),其中所有-反式维甲酸被运送到RPE,异构化为11-独联体异构体,然后运输回视网膜。第二种是一种不太为人所知的通路,位于视网膜内部,涉及Müller细胞和视锥内节段,被认为有助于视锥视觉色素的再生。
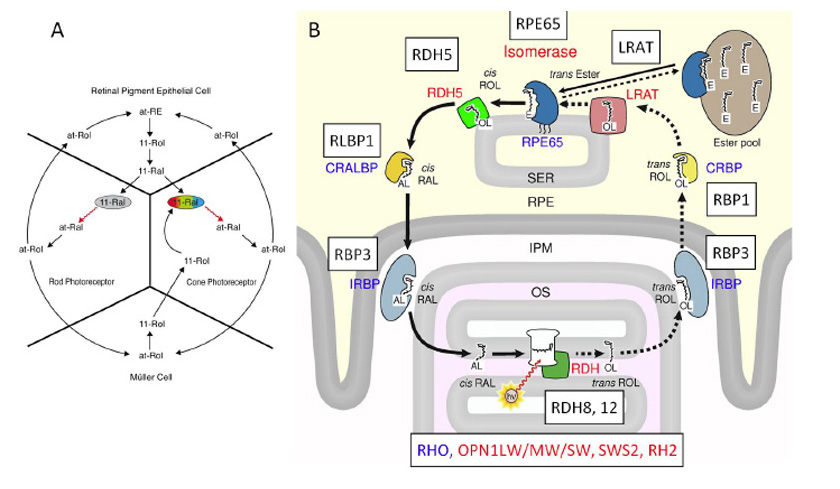
Intra-retina类维生素a周期
尽管在过去四十年中,大多数关于类视黄酮循环的研究都集中在RPE循环上,但锥体光感受器早于杆状光感受器这一事实提出了一种明显的可能性,即脊椎动物眼睛的祖先类视黄酮循环可能是包括视黄酮细胞和Müller细胞在内的视网膜内循环,RPE循环随后进化而来。视网膜内的循环似乎更简单,因为它涉及到所有-的异构化反式视黄醇(维生素A)直接到11-独联体视黄醇(图27A底部)通过Müller细胞中所谓的“异构酶2”,而不需要像RPE中那样进行初始酯化和随后的异构水解酶反应(见下文)。最近的一份报告表明,这第二种异构酶可能是去饱和酶,DEGS1(197),但其他有助于视网膜内循环的酶的身份才刚刚开始出现。不幸的是,由于对这个循环的研究还不够深入,还没有证明研究其进化根源是可能的。
RPE类维生素a周期
最近的两篇论文在确定经典RPE类维生素a循环的起源方面取得了重要进展。Albalat(198)和Poliakov等人(199)已经表明,这种循环只存在于脊椎动物中,因为头脊索动物和被囊动物不具备所需的酶,这意味着这种循环在图1中#4到#5的1亿年间隔中进化。
类视黄酮循环中的两种关键酶是LRAT(卵磷脂:视黄醇酰基转移酶)和RPE65。LRAT能酯化维生素A (all-反式视黄醇)-反式视黄醇酯,它是RPE65的底物,在所谓的“同分异构体水解酶”反应中,它实际上进行异构化,同时裂解酯键。这一步生成了11-独联体视黄醇随后被氧化成醛,11-独联体视网膜,然后转移到视网膜。
Albalat(198)进行了一次在网上在无脊椎动物中寻找类视黄酮加工的遗传机制,他分析了他发现的成分可能的功能。他的结论是:“基因组调查、系统发育重建和与脊椎动物类视黄周期相似的无脊椎动物成分(即Rdh8、Rdh12、Lrat、Rpe65、Rdh5、Rlbp1和Rbp3)的结构分析,并没有为脊椎动物以外的类视黄周期遗传机制的存在提供任何进化或功能支持。”
Poliakov等人(199)通过实验确定七鳃鳗和被膜中关键酶LRAT和RPE65(以及类似分子)的功能活性,扩展了这一方法。七鳃鳗(Petromyzon绿)的研究结果表明,LRAT和RPE65都是存在的,而且都具有功能,而且RPE65发挥同分异构体水解酶作用的关键位点与颌骨脊椎动物的位点非常相似。另一方面,他们的系统发育分析证实,RPE65和LRAT在头脊索动物(文昌鱼)或被膜动物(Ciona intestinalis).他们表明,一种酶以前被认为(并命名为)是一种CionaTakimoto等人(200)的RPE65的同源物没有同分异构体水解酶活性,证实了Kusakabe等人(9)的报道,他们表明它的功能是作为一种BCMO (β-胡萝卜素单加氧酶)。
总之,这些结果表明,从BCMO到真正的RPE65同分异构体水解酶的关键转变,以及LRAT的起源,都只发生在被囊动物分化后的脊椎动物谱系中(无颚和有颚)。因此,RPE类视黄循环只在脊椎动物中进化。
在许多无脊椎动物中,人们多年前就知道视紫红质可以在黑暗中再生(201),但关于这是如何发生的证据很少。在果蝇最近获得的证据表明,类视黄酮循环可能存在,尽管与RPE(202)中的证据有很大不同。
脊椎动物视网膜中类视黄循环起源的场景/RPE
根据我们目前对视网膜内和RPE类视黄酮周期的了解,我们可以假设脊椎动物眼睛中类视黄酮加工的进化过程如下:
J-1)大约在这个时候,脊索纤毛视蛋白失去了它们的双稳态(光可逆)特性,取而代之的是释放它们的全-反式类视黄醇,视网膜内的类视黄醇处理周期出现了,它利用Müller细胞中的异构酶(目前称为异构酶2)从其全-异构化视黄醇反式其11岁独联体的形式。
J-2)随后,大约在RPE成为视网膜对面的单分子层的时候,两种关键的酶LRAT和RPE65进化了,RPE能够在类视网膜再异构化中发挥额外的作用。
13视网膜在宏观水平上的发育
脊椎动物眼睛进化的线索可以通过研究其胚胎发育来获得,但需要谨慎。近200年前,卡尔·恩斯特·冯·贝尔(203)观察到,成年形态迥异的生物体,其胚胎彼此非常相似。他提出,胚胎所经过的发育阶段可能在某种程度上反映了生物体的进化史;von Baer的观点最近的评价,见Brauckmann(204)。
厄恩斯特·海克尔(Ernst Haeckel, 205)将这一概念扩展到“个体发育再现系统发育”的极端观点;也就是说,发展的每一步都反映了进化序列中相应的一步。这种严格的关系的问题是,它意味着进化只能通过改变或增加胚胎发生的最后阶段来实现,而这显然是不正确的。然而,尽管发育的所有阶段都受到进化的影响,但有一个明显的趋势是,更古老的步骤被保留下来,因此,在进化早期建立的基本身体结构似乎很少在胚胎发生过程中发生重大改变,相反,似乎有一个趋势是,新进化的特征被“附加”到胚胎发生的后期阶段。
因为发生在遥远过去的事件很可能发生在眼睛进化在眼睛的过程中有没有留下残迹的发展,现在我们将研究眼睛胚胎学的几个方面:首先,颌骨脊椎动物、七鳃鳗和盲鳗的眼睛发育的总体顺序;第二,脊椎动物视网膜细胞分类的顺序分化;第三,光感受器的成熟及其神经连接的形成。不可避免地,这导致了视网膜双极细胞的性质和起源。对这些主题的分析为视网膜的进化提供了重要的线索。
下颌脊椎动物间脑光敏器官的发育
在脊椎动物胚胎发育过程中,前神经板(视野)的一小块区域包含细胞,这些细胞将产生几种不同的光敏器官:成对的侧眼、松果体器官和松果体旁器官或顶眼,尽管并不是所有脊椎动物都存在(图28;(206, 207))。这些器官中的每一个都是发育中的间脑的外翻;背侧外翻开始是一个单一的凸起,但随后变得狭窄,分裂成成对的外翻,占据了吻侧和尾侧的位置,尾侧囊泡成为松果体器官,吻侧囊泡成为松果体旁器官或顶眼(206)。这些器官彼此具有密切的同源性,这些相似性的进化意义已在许多研究中得到强调,包括Reichenbach和Robinson (208), Vigh等人(207,209),Ekström和Meissl (210), Klein (211), Mano和Fukada(212)。
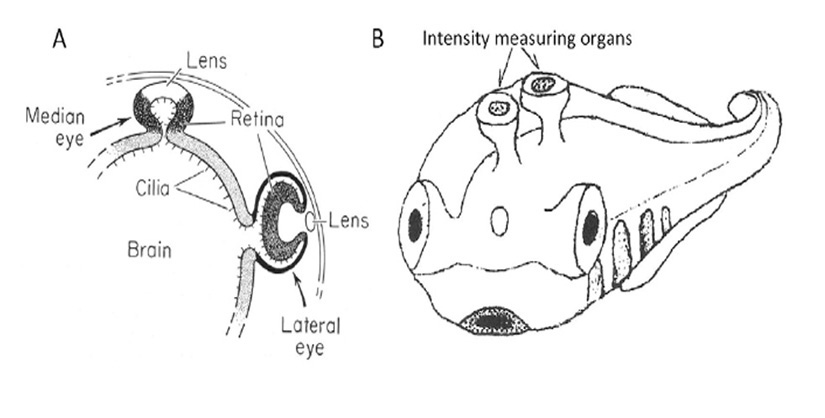
发育中的间脑囊泡的极性。从间脑(视网膜、松果体、松果体旁、顶叶)生长出来的正在发育的光敏生长物变得相当扁平——对比图10A中松果体的七鳃鳗和图29f中侧视视网膜和RPE。这两种器官之间的一个显著区别在于形成视网膜(或其同系物)的囊泡的一侧。在松果体和松果体旁器官以及顶叶眼中,主要是囊泡的近端(离大脑最近)发育成两层视网膜,因此光感受器的外节段(投射到管腔内)指向外部世界。由于投射神经元位于外表面,它们的轴突不需要穿透器官,可以直接进入中枢神经系统。器官的远端(面向外部世界)发育不同,在某些情况下为非光敏感组织;例如,在顶眼中作为所谓的“晶状体”,而在松果体中作为一层较薄的层,要么包含偶尔的光感受器,要么包含有时对紫外线敏感的光感受器(71)。与这些背侧器官相比,侧眼的极性是相反的,近侧发育为单层RPE细胞,远侧分化为多层视网膜。下面将提出,这种倒转极性的信号来自于茎类脊椎动物的一个祖先基板,该基板随后进化为晶状体基板。
在顶眼和非哺乳脊椎动物的松果体器官中,胶质细胞(光感受器嵌在其中)是着色的;在顶眼中,这些胶质细胞含有黑色色素颗粒(213),而在松果体器官中,它们含有反射细胞器,呈现板灰色外观。
宏观水平的侧眼发育。发育中的间脑的成对横向外翻产生了眼睛,其顺序(图29和下面的动画)由Walls(184)描述得很好,最近的Martinez-Morales & Wittbrodt(214)对此进行了综述。在吻侧神经板的每一侧形成一个凹陷,称为视神经槽(图29b)。随着神经板向上和向内折叠(图29b,c),汇合并闭合形成神经管,这些生长区域向外膨胀(图29d),现在称为视神经泡。然而,腔内空间似乎是最小的,因此囊泡的内表面保持有效的相互接触,有点像一个泄气的气球。膨胀的视囊泡和表面外胚层(图29e)之间的接触,在一个被称为晶状体基板的区域,引起了两者的变化,外胚层逐渐增厚,最终形成晶状体(图29g)。虽然有时有人认为视杯内陷是晶状体生长的物理结果,但事实并非如此,因为正常的眼杯和视网膜可以在晶状体缺失的情况下发育,如人类先天性原发性无晶状体(215)。值得注意的是,这种发育也可以发生在细胞培养中,在没有外胚层或晶状体组织的情况下(216)。
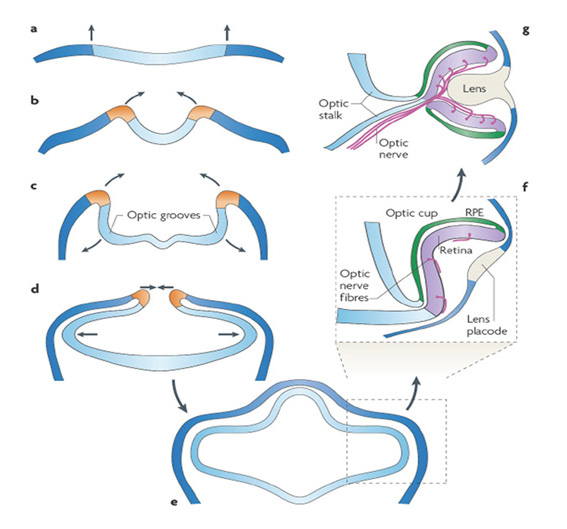
点击下方观看脊椎动物眼杯发展的动画:
兰姆等人(3)辅助动画S1
视网膜和RPE的描述。侧囊泡进入RPE和视网膜的初始极化受许多细胞外信号分子的控制,包括表面外胚层释放的fgf,但也涉及Bmp、Activin和Wnt。最近,有证据表明Pax6参与了一个正反馈电路,一旦RPE和视网膜的初始规范开始,视网膜/RPE内的信号就会自我强化(217)。这些作者提出,当Pax6与Mitf或Tfec一起表达时,它作为一种促rpe(即抗视网膜)因子,而与其他视网膜生成基因一起表达时,它作为一种促视网膜因子。因此,一旦偏向最初的视网膜/RPE极性,这种偏向就会通过发展中的视网膜/RPE内部的信号事件而自我强化。
一个更令人兴奋和有趣的最新发现是,当小鼠胚胎干细胞的聚集物完全在体外培养时,就像3D培养中的囊泡一样,它们能够自主组织成具有适当定向的视网膜和RPE(216)的眼杯,如下图所示。这一观察结果再次与视网膜/RPE内部信号系统的概念相一致,该信号系统可以在缺乏外部信号的情况下调节视网膜/RPE的发展。
点击下方观看眼杯体外自发形成的电影:
Eiraku et al (2011)补充信息,特别是电影2
细胞的协调迁移。在几项使用延时视频成像(218-220)的研究中,已经对斑马鱼眼泡形成的早期步骤进行了监测,因此可以看到单个细胞,并跟踪它们的迁移。这些研究的电影(下面链接)提供了令人印象深刻的斑马鱼细胞协调迁移的可视化效果。因此,宏观发育(例如,如图29所示)可以被可视化为单个细胞的协调迁移,首先主要是向外迁移,然后通过与基板接触位点附近的细胞向内迁移的组合,以及新细胞不断向外迁移进入囊泡(见电影)。
点击下方观看斑马鱼囊泡和眼罩形成过程中细胞迁移的影片:
英格兰等人(218)补充材料,特别是电影S1
雷伯德等人(219)支持在线材料,特别是电影S1
Kwan等人(220)补充材料,特别是电影S1
脊索间脑光敏结构进化的情景
我们似乎有理由认为,上述脊椎动物胚胎发育中的事件可能代表了脊索动物进化过程中的一系列步骤,即在被囊动物分化和脊椎动物出现之间的间隔(即图1中#4和#5之间)。因此,我们认为发生了以下步骤(也在图30的示意图中解释过):
K-1)早期脊索动物的前神经管包含一个光敏区,它将轴突发送到神经系统的相关部分(例如,发送到下丘脑的前部)。
K-2)内表面(与脑脊液接触)包含纤毛光感受器,而微绒毛光感受器(后来简单地变成投射神经元)位于外侧(图30A)。含有色素的细胞也存在。
这个感光区域扩大了,最初可能只是作为增加光感受器数量的一种手段,从而增加生物体捕捉光的能力。在此过程中,它从不同的方向向外隆起,分别是背侧和侧侧(图30B)。
K-4)随着这些区域的扩大,纤毛光感受器和微绒毛光感受器(即投射神经元)的体体保持紧密相连,而投射神经元的轴突延长,从而继续到达中枢神经系统的目标;因此,细胞体仍然紧密相连,但投射神经元的轴突“伸展”了。
每个鼓包的内部(管腔空间,包含脑脊液)保持相对较小,因此上皮细胞(包括纤毛光感受器和色素细胞)的内表面折叠在一起,像一个泄气的气球而不是一个膨胀的气球。
K-6)最初,这双层上皮组织在空间上可能是均匀的,纤毛光感受器、色素细胞和投射神经元分布在整个器官表面(图30未显示)。
K-7)在适当的时候,每个器官(背侧和侧侧)都倾向于发展成一种极性,其中一侧仍然主要感光,而另一侧则发挥不同的作用(例如作为顶眼的晶状体)。
K-8)在侧隆起中,而不是背隆起中,这种极化的方向是这样的,只有远端(外部)侧保持光敏,只有近端(更接近大脑)产生色素细胞(见图30B)。这样一来,纤毛光感受器就占据了入射光照射的整个截面积,色素细胞位于光感受器的后面,但又与光感受器保持着密切的接触。
在这些外侧器官中,轴突继续穿过外表面,现在包括视网膜的玻璃体表面。当它们向中央移动时,它们聚集成一束,沿着光学杆下的凹槽运行(图30C)。随着视网膜/RPE的持续横向向下扩张,视柄周围狭窄的间隙成为视裂。在进化的某个阶段(可能是后期),这个裂缝被封闭了,因此视神经穿透了外侧眼罩。
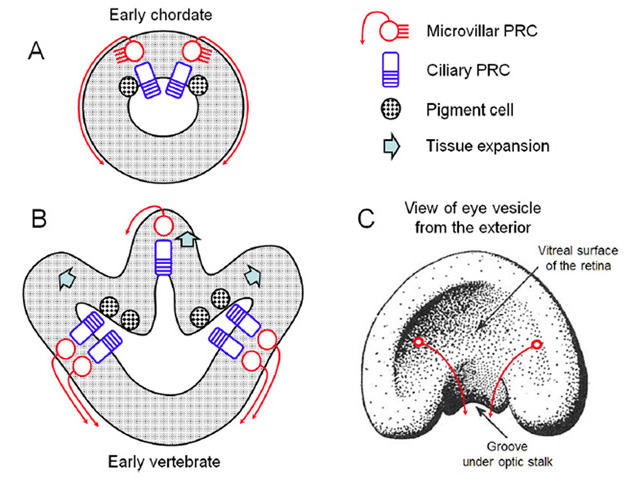
七鳃鳗和盲鳗的眼睛发育
七鳃鳗眼发展。七鳃鳗有一种幼虫(ammocoete,图13A),在蜕变成成鱼之前,它的发育要经过5年或更长时间(参见Dickson & Collard(84))。阿莫珂特实际上是失明的,它的眼睛和盲鳗的眼睛相似,因为它的眼睛很小,而且埋在皮肤下面。眼部发育的初始阶段发生在阿莫科动物的早期,与下颌脊椎动物眼部发育的初始阶段相似。然而,眼睛的发育就会受阻;在余下的ammocote阶段(例如,可能5年),眼睛的主要变化是,随着动物的生长,它(和它的晶状体)慢慢变大。
视网膜的大部分未分化,有较厚的神经母细胞层,但狭窄的中心区域(约50 μ m宽,靠近视神经)分化(84,221 -223)。虽然有报道称脊椎动物视网膜的所有常规层都存在,但重新检查这些显微照片表明,双极细胞实际上可能不存在于膜状视网膜中。一个缓慢的神经分化过程会持续数年,随着变态的临近,这个过程会加速。分化细胞的区域向外扩散,细胞按以下顺序分化:神经节细胞、无分泌细胞和水平细胞、光感受器,最后分化为双极细胞(21,222);即大致上与颌骨脊椎动物在胚胎发生过程中发生得更快的顺序相同(见第14节)。
然后,在变形时,七鳃鳗的眼睛迅速恢复发育。眼睛和晶状体迅速生长,视网膜分化完成,角膜分裂成巩膜层和真皮层(允许眼睛相对于表皮移动,就像颌鱼一样;(224)),眼外肌肉发育,眼睛在表面突起,成为脊椎动物式的视觉器官(图13B)。
八目鳗类鱼眼睛的发展。Price(225)首先研究了盲鳗胚胎的发育,Dean(226,227)和von Kupffer(228)也给出了全面的描述,尽管这些研究中只有有限的关于眼睛的数据。Allen(65)和Stockard(229)对盲鳗眼睛的胚胎发育进行了研究,特别是关于晶状体的起源(或缺失)。这些早期研究的综合结果与以下描述一致。
与胚胎的大小相比,发育中的眼睛总是非常小。在非常早期的阶段,视囊泡接触表面外胚层,可以看到一个明确的“晶状体基板”,尽管即使在其最厚的时候(在一个15毫米的胚胎中),这个基板也只有几个细胞深(229);此后晶状体基板变薄并消失。Allen(65)的结论是,如果晶状体基板的增厚代表了晶状体的发育,那么“我们应该有一个明确的发展受阻的案例,导致胚胎状态延续到成年生活。”这一观点得到了视网膜条件的有力支持,视网膜在视杯内外层之间有分隔空间,虹膜发育不全,脉络膜裂隙经常持续存在。”
在这些研究之后,在大约100年的时间里都没有证明获得可存活的受精卵是可能的;如果能使用仓谷茂实验室最近开发的方法继续那些早期的研究,那将是令人兴奋的。
回旋口眼发育对脊椎动物眼睛进化的意义
根据上面描述的环口和颌状脊椎动物眼睛发展的比较方面,我们推测了以下对脊椎动物眼睛进化的影响(3):
L-1)盲鳗眼睛的“发育受阻”(65)可能反映了盲鳗眼睛的发育未能超越脊椎动物眼睛进化的早期阶段。盲鳗不经历变态,在没有变态时发生的发育变化的情况下,也许盲鳗的眼睛在其早期的进化状态中被“冻结”了。换句话说,盲鳗的眼睛发育(也许是所有的发育)可能与一种稚嫩相对应。
L-2)如果这是正确的,那么盲鳗的眼睛和视网膜为了解脊椎动物眼睛进化的一个重要阶段提供了一个宝贵的窗口,并有以下附加的含义。
在进化的某个阶段,干脊椎动物的视网膜与盲鳗的视网膜相似,光感受器直接接触投射神经元(神经节细胞)。有关这一概念的进一步证据将在第14节视网膜连通性成熟中提出。
盲鳗的“晶状体”基板未能发育成晶状体,这表明该基板的祖先功能可能是影响视网膜/RPE的偏振方向(见上文),而不是形成晶状体。在此基础上,晶状体的进化可能发生在由折叠的两层视网膜组成的成对横向“眼杯”的进化之后。
L-5)同样,眼外肌在侧眼杯出现之前也不会进化。
综上所述,这些解释进一步表明,配对的外侧视网膜的原始功能并不是成像视觉,因为有证据表明,这些视网膜在晶状体、眼外肌或视网膜中除了最基本的计算能力以外的任何东西进化之前就已经存在了。相反,这些松果体器官的侧侧对应物可能在昼夜节律和/或季节计时方面,或者可能在阴影检测方面,甚至在控制轴向(即滚动)方面发挥作用。
因此,有人认为在脊椎动物进化的早期阶段,后“2R”基因组复制,但在圆口动物从颌骨谱系分化之前,在干脊椎动物中已经进化出一对带有RPE的原始外侧视网膜。这些视网膜拥有五种“视觉”睫状体视蛋白的早期形式,在视锥细胞和杆状光感受器中表达。双极细胞没有进化,也没有晶状体或眼外肌。据推测,“晶状体基板”的原始功能是通过释放成纤维细胞生长因子(fgf)等分子来指定外侧眼杯的极性。
有人认为,现存盲鳗的眼睛中保留着类似的状态,这可能是幼小的结果。七鳃鳗(以及它们的祖先)眼睛发育的后续阶段发生(可能已经发生)缓慢,主要是在变态过程中。但在现存的下颌脊椎动物中,眼睛发育的后续阶段发生得很快,与早期阶段密不可分。
14脊椎动物视网膜的分子特征与细胞分化
脊椎动物视网膜发育中神经元的分子特征
在原口动物和后口动物发育中的神经系统中,神经元细胞命运的规范似乎是由表达的转录因子(主要是bHLH和同源结构域超家族)的组合编码决定的。在分化细胞中,转录因子的特殊组合调节着控制细胞形态、轴突生长和效应分子(如转导级联、神经递质合成酶、离子通道等)表达的程序。关于转录因子在眼发育跨门的部署,见Vopalensky & Kozmik(6)。
为了深入了解组成细胞的进化史,Detlev Arendt和他的同事将构成细胞类型分化基础的分子因素的比较应用到视网膜上,包括生物内部和生物之间。(231)。阿伦特将这种方法称为“比较分子细胞生物学”。
姐妹细胞和同源细胞类型。Arendt(231)将“姐妹”和“同源”细胞类型定义如下。在生物体中,“姐妹细胞类型”是指通过细胞类型的多样化,从一个共同的前体进化而来的细胞类型。在进化树中,“同源细胞类型”是指从被比较群体的最后一个共同祖先中相同类型的前体细胞进化而来的细胞。利用这些定义,他认为,在任何脊椎动物物种中,水平细胞、无分泌细胞和神经节细胞是“姐妹”细胞类型,它们是从单一的祖先细胞类型演化而来的。在整个门中,他认为横纹肌光感受器和脊椎动物视网膜神经节细胞是“同源”细胞类型,从最后共同祖先(双侧)的微绒毛光感受器进化而来。
图31比较了脊椎动物视网膜不同细胞类型中表达的转录因子和效应基因,证明了前一种说法。锥状光感受器和杆状光感受器显然是姐妹细胞类型,由共同的纤毛光感受器演化而来(见第11节)。尽管Arendt对视网膜双极细胞的起源持模棱两可的态度,但随后将讨论双极细胞也是纤毛光感受器的姐妹细胞(第16节)。
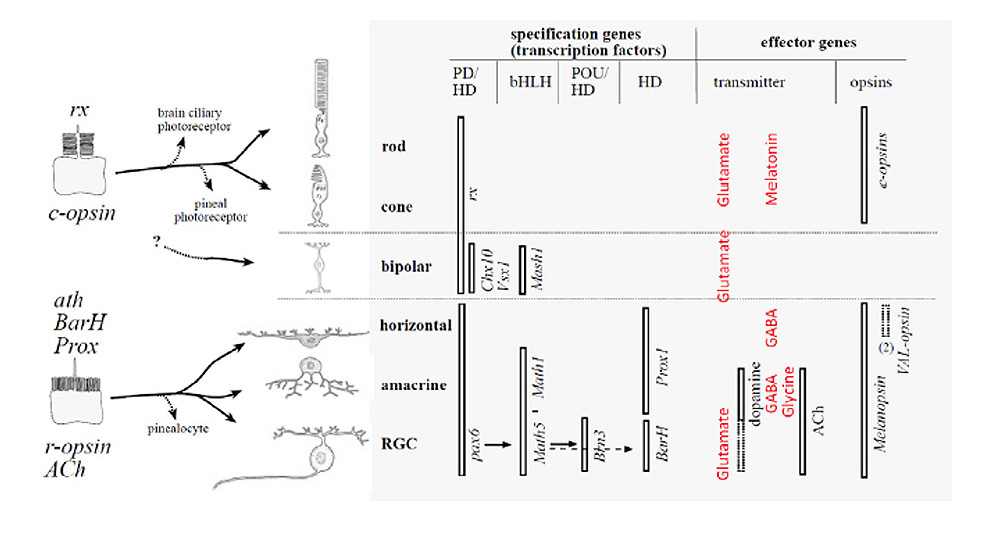
图31的下半部分显示,水平细胞、无分泌细胞和神经节细胞中表达的转录因子和效应基因非常相似(详细比较见Arendt(231))。此外,最显著的是在神经节细胞的情况下,这些因素与分化的基础因素和指定横纹肌光感受器的光转导的因素从根本上是相同的果蝇.根据Arendt的分析,水平细胞、无分泌细胞和神经节细胞似乎是从祖先的微绒毛光感受器进化而来的,所以它们不仅是“姐妹”细胞类型(彼此之间),而且它们也是原始口横纹肌光感受器的“同源”细胞类型。
并发症。Davies等人(94)指出了使这个相对简单的解释复杂化的两个观察结果。首先,尽管假设它们的共同起源是微绒毛光感受器,但一些水平细胞和神经节细胞表达c视蛋白(VA/VAL)。其次,这种c视蛋白VA/VAL似乎经常与r视蛋白黑视蛋白共表达。什么可能导致c视蛋白在祖先微绒毛光感受器后代的细胞中表达或共表达?除了视蛋白的这种复杂性外,现在已知的是转录因子的集合比图31所示的更广泛,因此细胞类型的分组不那么明显。
解决方案吗?解决这个难题(以及其他问题)的一个可能的解决方案是,认为所有脊椎动物视网膜细胞和RPE细胞都是从一种前双侧细胞类型进化而来的,这种细胞类型结合了光感受器细胞、色素细胞和胶质细胞的特征;例如,来自Arendt等人提出的“劳动分工”模型中设想的那种祖细胞(5)。这种观点似乎与脊椎动物视网膜细胞分化的知识一致(见下一小节),在该章节中,祖细胞最初在细胞命运中不受限制,并有可能形成任何类型的视网膜细胞。通过这种观点,我们就不需要限制所有脊椎动物视网膜/RPE细胞类型都经过严格的“睫状体光感受器类型”或严格的“微绒毛光感受器类型”的前因子才达到目前的形式。相反,一些细胞类型(如水平细胞)可能是从没有形成纤毛或微绒毛形式的光感受器的中间细胞类型演化而来的。如果是这样的话,那么视网膜和RPE中的现代细胞可能与祖先纤毛、微绒毛、色素或胶质类中的另一种具有相同的特征,这可能类似于一种混合搭配的方式。
在对脊椎动物视网膜细胞分化的简要描述之后,我们将继续阐述这一概念。
脊椎动物视网膜的细胞分化
关于视网膜细胞分化和神经元发育的综述,读者可参考Webvison的《视网膜神经发生》以及Sernagor等人(232)、Agathocleous和Harris(233)、Swaroop等人(234)、Reese(235)和Chen等人(21)的章节。这里给出了最简短的摘要。
视囊的神经上皮细胞将产生视网膜,它的细胞复制周期与大脑其他区域的细胞复制周期基本相同。祖细胞的体细胞体在神经上皮细胞的外表面(心室)和内表面(玻璃体)之间以一种特殊的模式垂直迁移,当子细胞退出细胞周期时,它的体细胞体就会迁移到视网膜层内的最终位置。分化开始于视网膜中央,向外延伸至视网膜外围。
命运,能力,和出生顺序。最初,祖细胞的命运不受限制,有能力形成任何类型的视网膜细胞。然而,不同种类的视网膜细胞“诞生”(即退出细胞周期)的顺序在一个物种中是很明确的,如图32所示,在脊椎动物物种中也很保守。特别是,大多数锥体光感受器在大多数杆状光感受器之前出生,而大多数双极细胞出生很晚,尽管在不同类别的生成中有时间重叠(236,237)。
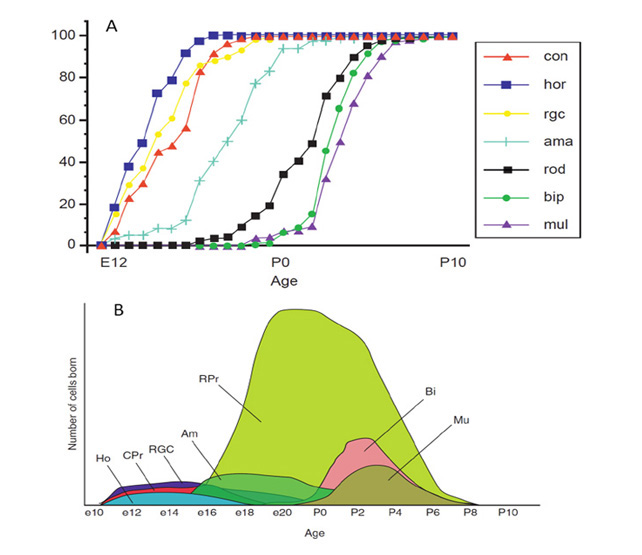
许多模型被提出来解释已经积累的视网膜细胞命运的实验结果,其中三个模型如图33所示。一个被称为序列能力的模型(图33A(a))提出,在任何时候,一个给定的祖细胞只具有产生一种或几种类型的细胞的能力,并且祖细胞经过这种类型的多种状态。另一个模型,渐进式限制(图33A(b)),提出早期祖细胞具有产生所有细胞命运的能力,而这种能力会随着时间的推移而逐渐丧失(限制)。
后一种模型下的显式方案如图33B所示,来自Reese(235)。该方案基于Wong & Rapaport(238)的研究结果,认为能力限制是祖代进步的单向序列。该图说明了(不寻常的)一个祖细胞产生整个视网膜细胞类别的情况。正如我们将在下面看到的,虽然不是每个分裂产生一个有丝分裂后的子细胞加上一个持续的祖细胞;有些分裂可以产生一对有丝分裂后的子细胞(例如一个神经节细胞和一个锥细胞,或一对水平细胞),而有些分裂可以产生一个Müller细胞和一个祖细胞,或一对祖细胞。然而,重要的是,在Wong和Rapaport(238)的方案中,在每一次分裂中,祖细胞的能力都逐渐受到限制,因此对应于“早期”阶段的细胞不再被创造出来。在图33B中,信号分子的假设影响由较宽的阴影箭头表示:对角线颜色的箭头表示来自不同类别新生细胞的信号,而垂直的灰色箭头表示Notch-Delta介导的侧抑制。
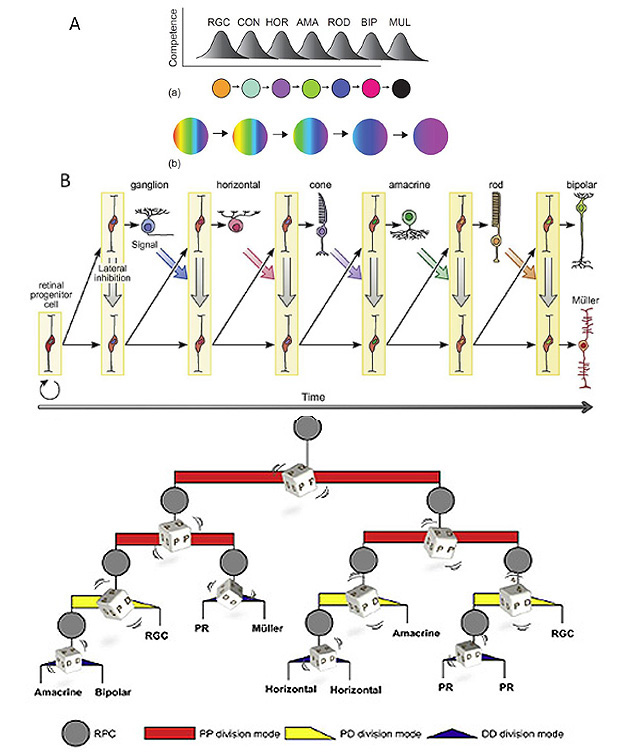
选择随机方案。最近,Gomes等人(239)提出了另一种随机模型(图33C),在该模型中,整个种群中细胞出生的顺序来自于每次细胞分裂时细胞命运的随机选择,他们和He等人(240)都获得了支持该随机模型的证据。直觉上,可能很难想象随机划分如何可能导致细胞类别的确定性组成,但He等人(240)提供了支持他们论点的数值模拟。
细胞分裂有三种模式(图33C)。在第一种模式中,称为“PP”,祖细胞产生一对祖细胞,导致祖细胞数量的增加。在第二种模式“PD”中,它产生一个持续的祖细胞和一个退出细胞周期的子细胞。在第三种模式中,它产生了两个子细胞“DD”,从而终止了谱系。He等(240)的分析表明,随着祖细胞进行多次有丝分裂,它们表现出从最初的首选PP模式到PD模式,最后到DD模式的转变。此外,子细胞的类型似乎与PD和DD分裂模式相关。特别是,大多数rgc产生于PD模式的划分;无分泌细胞产生于PD或DD模式;而水平细胞、光感受器和双极细胞多由DD分裂产生。他们还表明,无论是RGC的分化,还是PD分裂模式,都需要Ath5,这解释了为什么RGC的分化总是比其他类型的细胞更早。
通过这些(可能还有其他)影响个体随机分裂概率的因素的组合,宏观层面上的总体效果是视网膜细胞(平均而言)按定义明确的顺序诞生,同时视网膜细胞类型的最终组成也得到了适当的确定。
水平细胞。尽管人们普遍认为视网膜前体在外(心室)表面进行有丝分裂,子细胞垂直迁移到视网膜的最后位置,但水平细胞除外(至少在斑马鱼视网膜中是这样)。在斑马鱼水平细胞的情况下,绝大多数前体细胞(90%的最终种群)在成熟水平细胞的层流位置经历对称的“DD”有丝分裂模式,产生一对水平细胞子细胞(241)。这种在最终层流位置的分裂方式可能有助于水平细胞层的形成,并在这些细胞之间实现横向连接。
光感受器。光感受器的诞生和分化受大量转录因子和其他信使的影响,如Swaroop等人(234)所述。有趣的是,新生哺乳动物光感受器细胞的默认情况是,在没有其他影响的情况下,它将成为s锥(表达SWS1视蛋白)。在没有暴露在因子NRL/NR2E3的情况下,一般的光感受器将成为一个锥,而在缺乏因子TRβ2的情况下,它将成为一个s锥。将锥体暴露于TRβ2将导致其表达LWS视蛋白,(在人类中)在L-和M-(通过串联OPN1LW和OPN1MW基因)之间的选择受上游基因座控制区域的控制。将通用光感受器暴露于NRL/NR2E3将使细胞变成棒状。
如果假设祖先的球果类型是SWS品种,则产生s型球果的默认条件是有意义的。在脊索动物进化的阶段,在' 2R '之前,当这个祖先锥体和它的视蛋白复制时,需要一个额外的信号来指定另一种LWS命运。同样,在脊椎动物进化的后期阶段,当棒状体出现时,需要一个适当的信号来确定命运。
关于脊椎动物视网膜细胞进化起源的假说
根据前几节的观察,对脊椎动物视网膜/RPE细胞的进化起源提出了以下假设:
M-1)脊椎动物视网膜/RPE中的所有细胞类型都来自于一种多功能细胞的单一祖先类型,这种细胞存在于前双侧生物中,具有光感受器的功能,也含有屏蔽色素。
M-2)该细胞表达祖传视蛋白并激活g蛋白级联,其可能利用环核苷酸门控通道产生电反应;它也可能有附加的功能(例如运动)。
M-3)在进化过程中,两侧生物中随后的复制和分工产生了分离的纤毛和微绒毛光感受器,以及(在某些情况下)分离的色素细胞和分离的胶质细胞。
从古代起,光敏区的发育就涉及到多功能祖细胞的分裂。在早期的双侧动物中,该祖细胞可能只产生纤毛光感受器、微绒毛光感受器、色素细胞和胶质细胞。
M-5)在脊椎动物谱系中,可能在“2R”复制时间之后,但在颌骨和无颌骨谱系分化之前,其他细胞类型出现了,作为已有细胞的变体(包括微绒毛和纤毛光感受器)。在适当的时候,这些新细胞变成了水平细胞、无分泌细胞、杆状光感受器和两极细胞。
M-6)在扩张的脊椎动物侧囊泡中,极性出现了,可能是由来自祖先的“晶状体基板”的信号触发的,导致外层只产生上皮细胞(RPE),而内层产生所有其他类型的细胞(视网膜)。在缺少基板信号的背侧囊泡(松果体、松果体旁和顶叶)中,没有发生相同的极化,导致细胞种类的分布有所不同。
考虑到视网膜细胞生成顺序的数据,Dowling(242)提出,神经节细胞、视锥细胞和水平细胞的早期生成可能表明,这些是祖先视网膜中的原始细胞类别。他注意到水平细胞与神经胶质细胞有许多相同的特性,并认为最初的神经胶质细胞具有神经元的特性,因此成为水平细胞,并出现了一类新的神经胶质细胞(Müller细胞);随后加入无分泌细胞、杆状细胞和双极细胞。道林的方案为上述步骤M-5中水平细胞的起源提出了明确的建议。
分化与横向扩展的解耦.最后一点,值得注意的是,视网膜神经元的分化与眼球囊泡的侧向扩张并不是强制性耦合的。因此,Manns & Fritzsch(243)表明维甲酸对非洲爪蟾蜍胚胎可以阻止囊泡的形成,但在这种情况下,一组明显正常的视网膜细胞和RPE细胞可以在前脑背侧区分化,光感受器突出到第三脑室。
15光感受器到神经节细胞的突触传递
哺乳动物视网膜光感受器连接的发展
光感受器神经连接的发展过程是有趣的,可能代表了视网膜进化的一个信息遗迹。这一过程可能在哺乳动物物种中很常见,尽管在雪貂身上进行了最全面的研究(244,245),这被证明是很方便的,因为幼崽出生在视网膜发育相对不成熟的阶段,并且在之后的2周内不会睁眼。该过程由显微图和图34所示的图解顺序说明。
最初(在雪貂出生后的第一周)光感受器有一个简单的双极性形式,这使得仅从形态学的角度几乎不可能识别它们。但即使是这些非常年轻的细胞也会表达光感受器的特异性标记(如恢复蛋白、vGluT1、视紫红质),而且它们在表现出成熟光感受器的特征形态之前很久就会这样做。在图34A的免疫荧光图像中,由于对视紫红质的反应性,年轻的杆状细胞被标记为绿色,在杆状细胞外段分化之前,视紫红质分布在整个细胞质中。
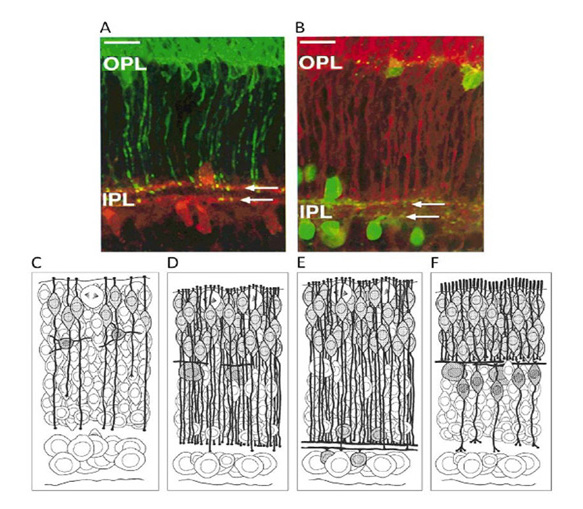
内丛状层的瞬态触点。在早期,每个未成熟的锥状或杆状感光细胞直接向内丛状层发送一个过程(图34C示意图)。在出生后的前两周,形成了越来越多的光感受器,它们的突起继续穿透新生的外丛状层的位置,尽管它们最终连接的水平细胞已经在它们最终的位置上,并开始向横向扩展它们的突起(图34D的示意)。在IPL中,年轻光感受器的过程终止于两个分离的亚层,与胆碱能无分泌细胞的分层过程一致(图34A、B的显微图和图34E的示意图)。光感受器末端表达突触蛋白synaptophysin和synaptotagmin,具有功能性突触的形态外观(245)。然而,尚未确定它们接触的突触后突起是无分泌细胞还是神经节细胞。
外层网状层的成熟触点。随后(雪貂出生后第14天左右),这些短暂过程从IPL收回,取而代之的是光感受器在发育中的OPL内进行突触接触(图34F示意图)。这一缩回阶段与双极细胞的成熟以及水平细胞和OPL的持续成熟同时发生。在OPL的发育过程中,每个光感受器末端形成一个带状突触,该突触首先与一对水平细胞突起形成二分体接触,随后与双极细胞突起接触,形成ON通路的成体三联体排列。几乎同时,各光感受器的远端突穿透外限膜,开始形成内段和外段。
上述的发展顺序,即信号从光感受器最初通过直接的突触接触到达输出层,但随后通过插入双极细胞进入传入通路而发生转变,这与视网膜进化过程中发生类似转变的概念是一致的。
除颌口视网膜外,其他器官的光感受器连通性
松果体器官:光感受器与神经节细胞的连接。在非哺乳动物脊椎动物中,松果体光感受细胞通过投射神经元(“神经节细胞”)上的带突触进行直接的突触接触,如图10A、D所示,并在Dodt(246)和Ekström & Meissl(247)中进行了综述。一般认为松果体器官只包含感光细胞、神经节细胞和胶质细胞,完全缺乏外侧视网膜的其他种类的神经元;然而,Ekström & Meissl(247)报道了一种推测的中间神经元表现出与光受体非常相似的电响应特性的单一病例。
松果体神经节细胞的反应主要是“光度OFF”单位,显示在黑暗中持续放电,并在任何波长的光照下抑制放电(69,74,246,248)。松果体光感受器和标准“光度OFF”松果体神经节细胞对中等亮度照明的典型反应如图12所示。对于次饱和强度,神经节细胞中的分级反应形式与光感受器中的反应形式非常相似。对于持续曝光,射速随强度的对数成比例减少,范围可达8个对数单位(74)。
此外,一些发光细胞是短暂的,在黑暗中显示出很少的峰值活动,但在光刺激停止时却会发出爆发力。偶尔也能看到颜色反应,这涉及到器官远端紫外线敏感的光感受器与主要的含视紫红质的光感受器之间的拮抗作用。
对这些结果的解释如下。在非哺乳脊椎动物的光敏松果体器官中,光感受器细胞通过带状突触连接直接与神经节细胞进行突触接触。黑暗中谷氨酸的高释放率导致神经节细胞持续的去极化和稳定的放电。在对光的反应中,光感受器超极化,减少谷氨酸的释放,并在神经节细胞中引起类似形式的分级反应。在神经节细胞中产生的符号保留反应相应地是“关闭”极性的;它们在稳定的光照下保持,并显示出与强度的对数关系。没有“ON”通路,也没有证据表明除了光感受器和神经节细胞外还有其他细胞参与。鉴于松果体器官可能早于脊椎动物的外侧视网膜,这些结果表明,在脊椎动物视网膜中信号流动的祖先通路可能是直接从视锥细胞到OFF视网膜神经节细胞。
盲鳗的“眼睛”:感光细胞与神经节细胞的连接。在盲鳗的眼睛中,唯一已知的神经元是纤毛光感受器和投射神经元。光感受器细胞的突触末端已被证明与突触后元件形成二分体接触,尽管迄今为止尚未确定这些突触后过程的身份;然而,我们假设它们是投射神经元。因此推测盲鳗的纤毛光感受器与投射神经元有直接的突触接触。
文昌鱼纤毛细胞与微绒毛细胞的接触?阿伦特和他的同事发现的这种分子同源性,存在于脊椎动物和果蝇视网膜,最近被扩展到文昌鱼的前眼。如第3节所述,Vopalensky等人(47)已经表明文文鱼额眼第1排睫状体光感受器与脊椎动物视网膜的锥体和杆状光感受器之间有密切的同源性;此外,第二行细胞可能与视网膜神经节细胞同源。在文昌鱼大脑的其他地方,微绒毛光感受器(约瑟夫细胞)直接与板层器官的细胞相邻,约瑟夫细胞“盖住”板层细胞(37,249,250)。虽然没有证据表明层状细胞实际上是光感受器,但这些报告至少表明,在进化过程中,微绒毛光感受器已经与层状纤毛细胞非常接近。
从光感受器到神经节细胞的突触传递的起源场景
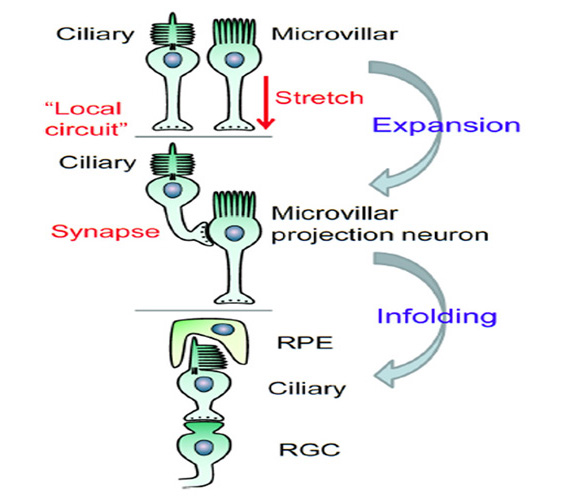
根据这些观察,我们提出了以下设想:在原始脊椎动物视网膜中,从纤毛光感受器到投射神经元的突触传递的起源(见图35):
N-1)在脊索动物进化的早期阶段,纤毛光感受器和微绒毛光感受器(由一个共同的祖先细胞演化而来)彼此相邻。它们对光分别有超偏光和去偏光反应。这些光感受器细胞嵌在色素细胞中,也可能嵌在分离的神经胶质细胞中,这些神经胶质细胞同样是从共同的祖先细胞演化而来的。
N-2)在这一阶段,纤毛光感受器通过“局部回路”或旁分泌机制传递信号,可能释放褪黑激素,也可能释放谷氨酸。这种旁分泌释放很可能具有昼夜节律或季节性的计时功能。
N-3)微绒毛光感受器通过轴突末端释放谷氨酸作为突触递质,将其输出信号发送到中枢神经系统的其他区域。
N-4)随着时间的推移,微绒毛光感受器也开始表达谷氨酸感受器,从而能够对来自纤毛光感受器的信号作出反应,并在适当的时候发展出成熟的突触接触。在此阶段,微绒毛光感受器作为投射神经元,可通过其轴突和突触输出快速传递纤毛光感受器的响应。
N-5)在某一阶段(可能与c视蛋白性能的提高和类视黄素在黑暗中再异构化的发明相一致),来自纤毛光感受器的信号提供了生物体所需的几乎所有信息,大多数投射神经元的光转导机制变得多余和凋谢。
N-6)这些细胞所在的区域是“视野”的前身,在未来会产生几个间脑光敏器官。它的扩展发生在第13节中。
16视网膜处理:双极细胞和光和暗通道
视网膜双极细胞的性质和起源
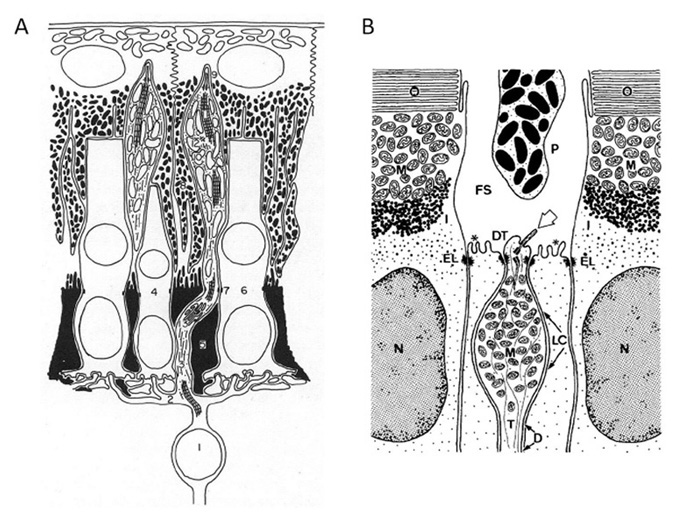
视网膜双极细胞与视锥和视杆感光细胞有许多共同的特征。在结构上,视网膜双极细胞具有相同的“双极”形状,最初表现为发展的光感受器,在其内部和外部节分化之前。
Landolt俱乐部。一些双极细胞有一个附属物,即Landolt’s club(图36),它与光感受器的内段惊人地相似(251-253)。该细胞器具有非运动纤毛的9+0型结构特征的微管,位于外膜,但与内膜不同的是它不产生外膜。Quesada和Genis-Galvez(254)在鸡视网膜中报道,在胚胎早期,所有的双极细胞通过Landolt 's club连接到外限制膜,在发育过程中,许多这些过程收缩。
转导。就转导机制而言,ON双极细胞的g蛋白级联与脊椎动物光感受器的光转导级联具有显著的相似性。此外,双极细胞的输出突触与光感受器的输出突触非常相似,使用的突触带结构在任何其他类型的视网膜神经元中都没有发现。最后,视网膜双极细胞中表达的许多蛋白质要么与锥体和杆状光感受器中发现的蛋白质相同,要么代表它们的异构体;例如,恢复蛋白、钾离子通道和带状突触的分子机制。
鉴于这些密切的相似性,以及视网膜细胞的分化顺序(图33),有可能将视网膜双极细胞视为视网膜纤毛光感受器的可能“后代”,从这个意义上说,它们似乎是作为已建立的纤毛光感受器主题的一种变体而进化的,因此继承了这些光感受器的许多特性。如果情况确实如此,那么人们可能会预期锥ON型双极细胞可能是从锥细胞进化而来的,而杆状双极细胞可能是从杆状而不是锥型双极细胞进化而来的。
另一种可能解释这些观察结果的可能性(虽然可能不太可能)是,光感受器和双极细胞是“原神经元”的独立后代,“原神经元”被假设为最古老的脊椎动物神经细胞类型(208,209,255)。然而,这一假设似乎与锥状光感受器和杆状光感受器是由祖先纤毛光感受器进化而来的证据不符。现存的一个被认为是祖先原始神经元的代表是文昌鱼、盲鳗(256)和七鳃鳗(257)以及下颌脊椎动物的中枢神经系统中存在的感觉“脑脊液(CSF)接触神经元”。
光信号通路通过双极细胞的进化场景
根据第14节中关于胚胎视网膜中光感受器接触发展的观察,以及上述关于双极细胞性质的观察,以及第14节中关于视网膜发育时间的发现,以下是在脊椎动物视网膜传入通路进化过程中可能发生的事件顺序:
O-1)在眼睛进化的阶段,囊泡已经向外侧凸起,形成了原始的视网膜/RPE,光感受器已经直接与神经节细胞形成突触接触,神经节细胞将轴突投射到中枢神经系统。
O-2)水平细胞进化,并以二分体结构在带状突触上与光感受器形成突触接触;它们提供了光信号的空间整合,并将这种空间平均信号反馈(反向)到光感受器终端,从而产生空间对比元素。
随后,进化出一种改良的“伪光感受器”细胞,它失去了表达视觉色素和一个外部片段的能力,但却表达代谢性谷氨酸感受器。
O-4)在视网膜发育过程中,这类新的细胞,视网膜ON双极细胞,将其树突末端插入光感受器双极细胞中,形成三联,而它的传入过程与它的前身(光感受器)经过相同的旅程到达IPL。
O-5)该细胞的一个变体表达离子化谷氨酸受体,成为OFF双极细胞。
总的来说,一套新的双神经元件被整合到原始的两层视网膜中,极大地扩展了视网膜执行时空处理计算的能力。
杆状信号的专用scotopic通路
哺乳动物的杆状通路中出人意料的视网膜回路的存在最早由Kolb & Famiglietti(258)、Nelson(259)和Kolb and Nelson(260)描述;参见“杆状信号的电路”,“所有无分泌细胞”和“双极细胞通路”的网络视觉章节。这一回路的细节被Strettoi等人(261)和Vaney等人(262)进一步扩展,不久之后,Strettoi等人(263)提出了哺乳动物scotopic(杆驱动)视网膜通路“背着”预先存在的光topic(锥驱动)通路的概念。巧合的是,就在这一年,Okano等人(115)发现杆状视蛋白由锥视蛋白进化而来。
就功能而言,这种背负式排列的主要优势在于,双锥加杆系统不需要在视网膜中重复神经处理通路,也不需要在大脑中重复一组神经纤维。相反,杆信号进入预先存在的复杂的锥神经处理电路,在那里它们可以无缝集成。对于视觉感知来说,这意味着一个人几乎不可能区分它是自己的scotopic系统还是phototopic系统正在被利用。
将杆信号注入哺乳动物锥体系统的方式看起来很复杂(图37),但它巧妙地避免了引入重大的折衷,特别是通过避免产生额外的噪声。从中视(黄昏)光水平到高暗视(月光)光水平,杆状信号通过缝隙连接传递到锥(图37中表示为ON2和OFF2),从而完整地利用了光通路。但在非常低的星光水平,一个单独的专用系统开始运作,利用棒双极电池和全无分泌电池。
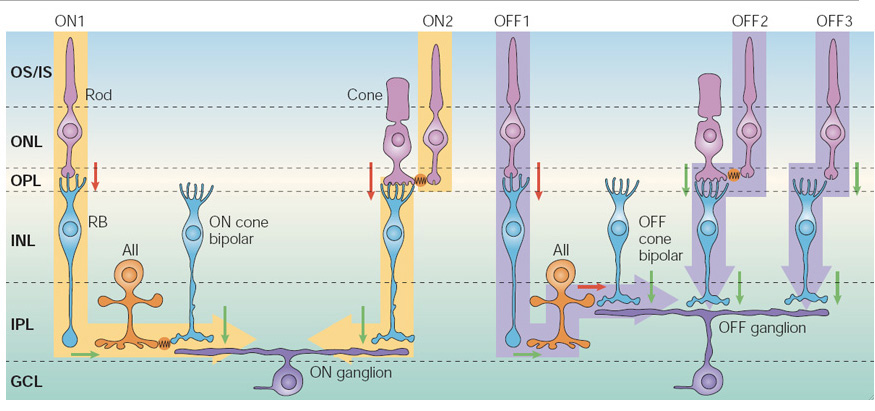
对于ON系统,如图37左侧所示,光致通路使用一个信号反转突触(红色箭头)从锥状光感受器到锥状ON双极细胞(通过代谢性谷氨酸受体机制),然后使用一个信号保留突触(绿色)从锥状ON双极细胞到ON神经节细胞。因此,在低光水平下,当视锥细胞对信号的贡献很小时,ON双极细胞是超极化的,而ON双极细胞到神经节细胞的突触是静止的。在较低的scotopic水平(指示ON1),杆状细胞的活性通过标记反转突触传递到杆状双极细胞,然后通过标记保留突触传递到全无侧侧细胞,因此昏暗的scotopic光照导致全无侧侧细胞的去极化(即与锥体ON双极细胞中任何受光刺激的活性的极性相同)。然后,在无附加噪声的情况下,通过间隙结电连接从所有的无分泌细胞连接到锥ON双极细胞(橙色表示),将闪点信号耦合到锥通路中。在强光下,当电极饱和,电极双极细胞和所有无分泌细胞被强烈去极化时,间隙连接可能会关闭,以防止不必要的信号的侵入。相反,如果杆状双极细胞通过突触直接连接到ON神经节细胞上(就像锥体双极细胞一样),一个主要的缺点就是当杆状双极细胞饱和时,会向光致系统注入大量的突触噪声。
对于OFF系统,如图37右侧所示,光致通路使用一个信号保存突触(绿色箭头)从锥到OFF双极细胞(通过离子化谷氨酸受体机制),然后第二个信号保存突触(绿色)从锥OFF双极细胞到OFF神经节细胞。结果,在静止黑暗状态下,锥体OFF双极细胞去极化,并不断以高速率向OFF神经节细胞释放突触递质(谷氨酸)。那么,在这种持续的突触输入存在的情况下,scotopic系统的活动是如何被注入到锥体OFF通路的呢?已被采用的答案是将全无分泌细胞的信号反转突触输入到锥体OFF双极细胞的突触前端(标记为OFF1)。通过这种方式,昏暗的射灯导致锥OFF双极细胞的突触末端超极化,从而抑制了突触递质持续的高速率释放,就像来自锥细胞的信号一样。
这一分析表明,在哺乳动物出现的时候,进化已经找到了一种方法,通过这种方法,将最近进化的杆的单光子探测能力叠加到已有的锥神经处理路径上,以无缝的方式,使用最小的额外布线,以一种方式引入最小的额外噪声,最小地干扰锥神经电路复杂的信号处理功能。然而,目前还不清楚这是否是早期脊椎动物视网膜所采用的第一种解决方案。
因此,发现除了哺乳动物以外的脊椎动物的视网膜中是否也有类似的全无分泌细胞回路是很重要的,但到目前为止,对这个问题似乎还没有确定的答案。在非哺乳动物的脊椎动物视网膜中,很明显,暗缩信号是通过高灵敏度的双极细胞处理的,其反应特性与哺乳动物的杆状双极细胞非常相似;例如,角鲨的Ashmore & Falk(264)。同样,七鳃鳗视网膜的ERG记录也显示出敏感b然而,杆信号和锥信号结合的电路还不完全清楚。一种可能性是,在非哺乳动物视网膜中,视杆和视锥信号仅在外丛状层水平上合并,通过视杆和视锥之间的间隙连接,也可能通过视杆和视锥之间的突触输入到单一的双极细胞。在这种情况下,双极细胞将是杆锥混合型,然后信号将通过传统的锥通路处理,包括双极细胞到神经节细胞。另一种可能是,视网膜内部可能存在类似于所有无分泌细胞的电路。在非哺乳动物脊椎动物的视通回路的细节被阐明之前,可能无法确定祖先脊椎动物的视网膜在多大程度上利用外丛状膜层和内丛状膜层将其杆状源性信号注入锥体通路。但无论在哪种情况下,很明显,作为最小值,已经存在的锥体系统的输出(神经节细胞)阶段被利用了。
在这个阶段,还没有足够的信息来证明在专门的scotopic通路的进化中有明确的步骤。
17空间(成像)视觉是什么时候在脊索动物谱系中出现的?
在前面的第13节中,我们讨论过,脊椎动物侧眼的视网膜最初是作为一种非成像的光敏器官进化而来的,与现存的非哺乳脊椎动物的背侧器官(松果体、松果体旁和顶骨眼)的两层视网膜有许多相似之处。但是双层视网膜是什么时候第一次出现的呢?那对“外侧松果体样器官”是什么时候,通过什么方式进化成视觉系统的成像器官的?beplay体育公司
没有证据表明,我们与被囊动物共有的最后一个共同祖先(图1中的#4)可能拥有类似视网膜的东西。另一方面,我们与七鳃鳗的最后一个共同祖先(图1中的第5点)似乎几乎可以肯定拥有一个完全成熟的脊椎动物式的眼睛,有三层视网膜、晶状体、眼外肌肉等。不幸的是,在那段时间里,从我们自己的谱系(图1中红色曲线所示)中分离出来的无数生物没有一个存活到今天。从化石记录中可以知道,在这段时间里分化出了几种灭绝的物种。尽管据报道,这些化石物种拥有“眼睛”,但它们的软组织保存非常不完善,甚至不知道这些器官是否含有晶状体。被称为“眼睛”的侧印化石似乎是黑色的,或至少是深色的,这可能与它们含有色素一致。
因此,目前似乎不可能更具体地说,外侧两层视网膜在图1第4点之后的某个时间出现,而成熟的外侧眼睛在图1第5点之前的某个时间出现。根据Erwin等人(20)提出的日期,这些差异分别发生在600和500年。因此,不幸的是,我们似乎无法更精确地把这些重要特征的相关演化时间限定在1亿年之内。
如果有可能精确地确定“2R”对全基因组复制发生的时间,那么就有可能缩小眼睛进化中感兴趣事件的时间范围。因此,纤毛视蛋白的四倍复制直到“2R”之后才出现,因此目前的五类纤毛视蛋白不可能在“2R”之前出现,因此(例如)视杆蛋白不可能在“2R”之前出现。
尼尔森和佩尔格(265)曾计算过透镜进化所需的时间。这一计算假设了一个功能齐全的视网膜已经存在,并且已经与大脑进行了适当的连接,但这一计算没有考虑到可能需要的其他变化,比如用来调节眼睛方向的肌肉系统的出现。因此,他们计算出的接近100万年的时间,似乎大大低估了将少量光感受器转化为一只功能性眼睛所需的时间。
相反,我们可以预期上面描述的转换可能会花费更长的时间,因为每个转换都必须涉及到一个很长的变更序列。将少量纤毛和微绒毛光感受器转化为松果体样视网膜需要许多步骤。此后,要添加双极细胞,添加晶状体和角膜,添加眼外肌肉,以及(可能是最需要的)在大脑中创建一个处理空间信息的处理中心,都需要大量的平行步骤。考虑到实现这些主要转变(首先是不可成像的视网膜,然后是成像的眼睛)所花的时间可能占据了目前认为可以利用的1亿年间隔的很大一部分,这似乎不是不合理的。
18结论:光感受器和脊椎动物视网膜的进化场景
下面的小节使用图1中的纪元编号方案,为光感受器细胞和脊椎动物视网膜的进化所涉及的众多步骤中更重要的步骤的顺序提出了一个设想场景。
早期后生动物:在第一点之前(>700 Mya)
感觉细胞。所有动物光感受器都是由一个g蛋白偶联受体(GPCR)进化而来的,它触发了一个g蛋白级联,这个级联很可能使用了环核苷酸门控通道(cngc)。
视蛋白。最初的视蛋白是在相对较短的时间(约1000万年)中产生的,就在700万年之前(见图2),在我们自己的谱系中,首先是平胎动物,然后是刺胞动物的分化之间。在此之前,GPCR的突变改变了它的结合特异性,使激活它的配体变成了视黄醛,修饰后的GPCR因此成为了视黄醛受体;与其他GPCRs一样,配体以非共价结合。(这种GPCR的近亲之一似乎引起了脊椎动物褪黑激素受体的产生。)在平胎虫分化后,这个视黄醛受体的突变使它的第7个跨膜螺旋(对应于K296)中有赖氨酸残留,因此视黄醛配体现在可以通过席夫碱键共价结合。进一步的突变在附近添加了一个带负电荷的残留物(对应于E181),充当反离子,使希夫碱被质子化。这种分子因此成为祖先视蛋白。它会在光谱的短波端被吸收,尽管我们不能确定它的峰值是在我们所说的光谱的“可见”部分还是在紫外线中。随后,在刺丝细胞分化之前,这种祖先视蛋白复制了两次,产生了视蛋白的三个主要分支,即C-、R-和RGR/ go -视蛋白。
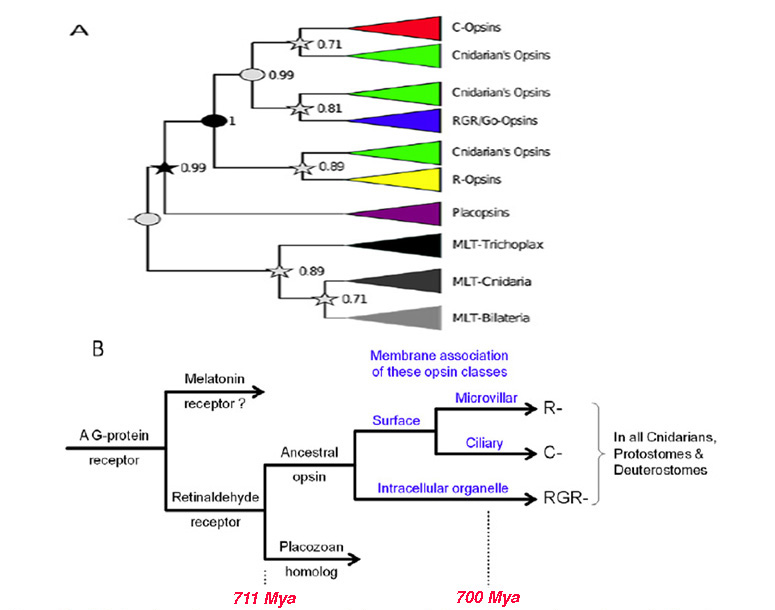
光感受器细胞和光转导级联。一旦祖先视蛋白出现,祖先的光感受器就诞生了。这一最初的光感受器细胞似乎与纤毛类细胞相似,视蛋白在纤毛膜中表达,并通过Gi和/或Gs级联调节环核苷酸水平和CNGC活性。这种原始的光受体细胞有可能具有多个g蛋白信号级联的成分,但目前所能获得的信息太少,无法确定其转导级联。在刺丝细胞分化之前的短时间内,可能已经存在两类含有视蛋白的细胞,一类在其纤毛表面膜表达祖先的c -视蛋白,另一类在胞内细胞器膜表达RGR/ go -视蛋白。祖传的RGR/ go -视蛋白的一个可能功能是作为一种视黄酮光异构酶,利用光转化全-反式11-的同分异构体独联体同分异构体。虽然使用r -视蛋白的独立的微绒毛光感受器可能也出现过,但它们后来在刺丝动物中消失了,这表明微绒毛类光感受器只在双侧动物中出现。
祖先光敏细胞的其他成分。这些早期后生动物的祖先光敏细胞很可能是多功能的,此外,它们还能表达可作为光屏障的色素,也有可能这些细胞具有运动功能(79)。拥有这些细胞的动物会非常小,没有可识别的神经系统。它们的细胞可以通过扩散的化学信使相互传递信号;即通过旁分泌释放。控制这些细胞分化的基本特征的化学信息传递系统需要在这个时候建立起来。因此,光敏组织特异性转录因子的祖先系统可能已经存在。
劳动分工。正如Arendt等人(5)所提出的,基因复制和进化的过程似乎导致了更专门化的子细胞的出现,这些子细胞之间分担了原本由单个细胞执行的任务。如果如上文所述,一个古老的多功能光感受器细胞表达了不止一类视蛋白和转导级联,还具有色素细胞的功能和运动作用,那么随着时间的推移,这些功能可能被委派给单独的专门细胞,这些细胞只继承了与原始功能的一个子集相关的成分。因此,运动细胞和色素细胞从进行光转导的细胞中分离出来;同样地,“光异构酶”视蛋白可以被分解到单独的细胞中(例如,色素细胞)。类似地,不同的视蛋白及其转导级联被分离成不同的细胞类型。
双侧口动物,原口动物和早期后口动物
早期的两侧对称。在双侧动物谱系中(即在刺丝细胞分化后),至少存在两种不同的光感受器细胞:表达c视蛋白的纤毛光感受器类和表达r视蛋白的微绒毛光感受器类。这两类细胞似乎有可能被用于不同的功能。一种可能是纤毛光感受器在昼夜和季节的计时功能中起作用,而微绒毛光感受器在趋光行为中起作用,为生物体提供趋光或避光的信号(例如在水柱中向上或向下)。
原肢类。随着原口口和后口口的分裂(在图1的#2处),一些原口口在少量的微绒毛光感受器前进化出一个晶状体,这可能成为一个眼珠或一个小网膜。也许进化出的信号导致了大量相邻小眼的复制/增殖,导致了复眼的形成。在后期,一些原气孔系的微绒毛光感受器发展出高度组织的亚细胞结构(横纹肌)和高度特化的转导级联,并产生横纹肌光感受器。在这些特殊谱系中出现成熟的横纹肌光感受器之前,微绒毛光感受器可能不是非常敏感是完全合理的。例如,在脊索动物谱系中,文昌鱼的微绒毛光感受器的敏感性远远低于它们进化出横纹肌结构的原始口体(46)。
早期的后口动物。在后口动物谱系中似乎没有出现这样的发育(以晶状体或横纹肌结构的形式)。因此,尽管微绒毛光感受器存在于后口,但它们仍然是相对简单的细胞,分布稀疏,有时与色素细胞结合。同样,在早期后口动物谱系中,纤毛光感受器的变化似乎相对较少。
脊索动物-早期事件:从第3点开始(约650万年以来)
C-opsins细化。从原始的c视蛋白到脊椎动物视蛋白的转变似乎涉及到一个漫长的过程。最重要的变化之一是反离子位点的重新定位(从181位到113位),因为这允许元II状态以一种实现更高的g蛋白激活率的方式被修改,也允许元II状态相当迅速地衰减,释放所有-反式视网膜。
在c -视蛋白的系统发育树中(图19),有几种现存的视蛋白存在于最基础的c -视蛋白和锥视蛋白之间,包括Opn3、壁视蛋白、副视蛋白、VA/ val -视蛋白,可能还有视蛋白。这些视蛋白的功能属性也趋向于遵循一个序列(章节5),似乎很明显,c -视蛋白的性能在脊索动物进化过程中稳步增强,在第一个锥视蛋白出现时达到一个高水平(章节5)。之后,这个祖先锥视蛋白复制形成SWS和LWS视蛋白。有被膜的的Ciona intestinalis,在第4点分裂(图1),拥有一个单一的c视蛋白Ci-Opsin1,它在VA/VAL附近演化(尽管这在图19中没有明确显示。因此,最终形成锥视蛋白的脊索c视蛋白的细化过程在被膜分化的时候只进行了一半,也许整个细化过程可能在图1中3到5的1.5亿年的大部分时间里持续了。
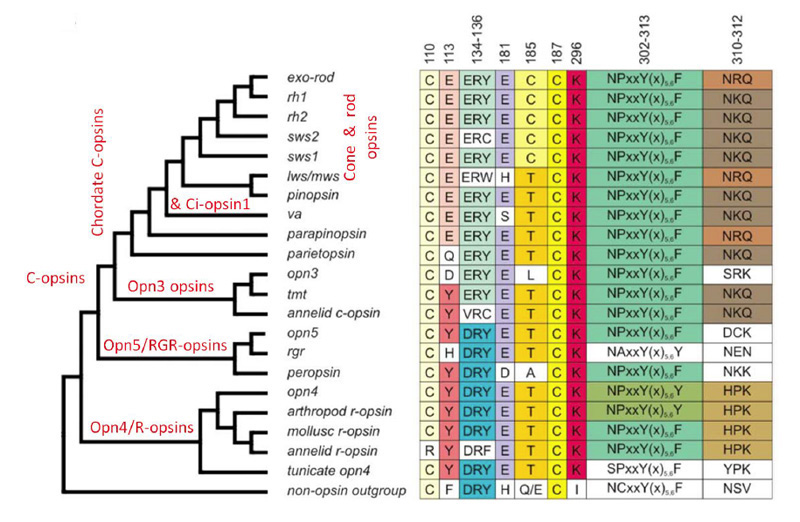
光转导级联的改进。在脊索动物的进化过程中,光转导级联似乎也发生了改进。这些变化包括:g蛋白α亚基(即转导素)的Gt变体的出现,显然是通过共同的Gαi / Gαt祖先的复制;cGMP特异性PDE6的出现;其调控型PDEγ亚基的出现;等等。虽然这些发展的确切顺序仍然不清楚,而且事件链无疑是漫长的,但似乎很清楚的是,在“2R”全基因组复制之前,祖先锥体光感受器中的光转导级联已经达到了一种状态,从本质上与现代脊椎动物锥体的状态相似。
纤毛光感受器到微绒毛光感受器的突触接触。很少有标志物表明纤毛光感受器和它们的微绒毛对应体之间的突触接触是在什么时候建立的(大多数微绒毛随后失去了对光的敏感性)。文昌鱼拥有紧挨着板层器官细胞的微绒毛光感受器(约瑟夫细胞),微绒毛细胞“盖住”层状细胞(第15节),尽管这些层状细胞通常被认为是纤毛光感受器,但这尚未通过电生理记录得到证实,也未在其中发现视蛋白。最近研究表明,在文昌鱼的“额眼”中,纤毛光感受器(第1排细胞)紧邻投射神经元(第2排细胞),投射神经元可能与视网膜神经节细胞同源(6)。因此,我们与文昌鱼的最后一个共同祖先可能已经有纤毛光感受器与微绒毛光感受器进行突触接触。
脊索动物向脊椎动物的转变:间隔#4 - #5(约600 - 500米亚)
在脊索动物谱系中,区分“眼斑”和“眼睛”的大多数转变似乎都可能发生在第4到第5的1亿年间(图1),大约在600-500米亚。在光敏组织本身,这些转变包括:大量扩大组织形成视网膜;一种类视黄酮在黑暗中再异构化方法的发明;视网膜处理能力的阐述,包括双极细胞的发明;一类锥体细胞转变成棒状细胞。在视网膜之外,这些转变包括:辅助结构的形成,包括晶状体、角膜、虹膜、眼内和眼外肌肉,以及处理视觉信息和控制运动系统的大脑区域的形成。
间脑光敏区扩大。在脊索动物进化的某个阶段,组成感光组织的区域在背侧和外侧都扩大了;大部分的扩张似乎发生在被囊分化之后。扩展的最初驱动力可能只是为了实现更高的灵敏度,这可能导致部署更大的区域来吸收入射光。一个有利于横向扩张的因素可能是伴随原始大脑的扩张和软骨头盖骨形成的下行光的遮挡。这种生物的优势不仅在于提高了对光的敏感度,而且还具有探测阴影的潜力和粗略比较入射光的方向。
在发生的组织扩张中,似乎只有微绒毛光感受器细胞将轴突延伸到它们在更中央的大脑区域预先存在的目标,而纤毛光感受器细胞继续以纯局部的方式发出信号。在某个阶段,它们的信号机制变成了突触,而不是旁分泌,尽管纤毛光感受器至今仍保留着释放褪黑激素的能力。
凸出的区域似乎有最小的腔空间,因此其相对的腔(内)表面彼此接触;每侧最内表面均有睫状体光感受器嵌于色素细胞和胶质细胞之间。这个不断膨胀的囊泡形成了一种“极性”,使两边变得不同,并在适当的时候产生了“视网膜”在一边和“RPE”在另一边的完全区别。这种极性发展的信号可能是由后来演化为透镜基板的祖先基板产生的。由于极化,面向外部世界的一侧不再形成色素细胞,而神经元件只在这一侧形成。这为生物体提供了一个主要的优势,就增加了对光的敏感性而言,因为器官的整个横截面区域现在都布满了感光器,而以前该区域的很大一部分被色素细胞所占据;相反,色素细胞现在被有效地放置在感光细胞的后面(就入射光的路径而言),但仍然与它们有物理接触。
这种扩张和鼓胀,结合最小腔体积的保留和器官极性的发展,是“由内而外”脊椎动物视网膜的产生原因。
在脊索动物进化的这一阶段,有人认为,原始的“眼睛”还没有进化出光学部件(晶状体等),或复杂的神经处理能力(双极细胞等),或定向运动系统,相反,它可能与现存盲鳗的“眼睛”非常相似。
类维生素a的黑色再异构化。伴随着在脊索c视蛋白中g蛋白活化效率的提高,活性后视紫红质发生光逆转至11-的能力的丧失独联体同分异构体形式(第5节)。这就需要释放all-反式类维生素a及其通过其他途径的再异构化。潜在地,这可能是利用了一种单独的光异构酶,沿着鱿鱼眼睛中使用的视网膜色素的方向——可能这种异构酶在某个阶段被使用过,因为RPE中存在rgr视蛋白,它能够这样做反式- - - - - - - - -独联体光异构化。然而,在脊索动物谱系中,更重要的似乎是在黑暗中异构化的化学路线(第12节)。在现存的脊椎动物物种中,有明确的证据表明视网膜内存在视锥细胞可获得的类视黄酮循环,尽管细节尚不清楚,起源也不清楚——但完全有可能,这种视网膜内循环可能代表了all-异构化的祖先途径反式形成11-独联体在黑暗中异构体。从RPE65或LRAT的缺失来判断,更广为人知的途径包括类维生素a运输到RPE和返回,是在被膜分化后进化的Ciona.
纤毛光感受器的胜利。在黑暗中进行再异构化的化学途径的出现很可能使脊索纤毛光感受器比利用双稳定光颜料的微绒毛光感受器具有主要优势。当动物从一个光线明亮的环境移动到一个光线昏暗的环境时,微绒毛光感受器就会被大量稳定的后视紫红质所负担,无法迅速将其转换回视紫红质;因此有用的视紫红质的含量会保持在很低的水平,而这些感光体的敏感度会长期下降。相反,纤毛光感受器可以通过释放all-来“适应黑暗”反式类维生素a和11-的再结合独联体视网膜形成视紫红质;为了长期保持这一能力,释放了一切——反式类维生素a需要化学异构化回到11-独联体在黑暗中。因此,当脊椎动物之前的脊索动物在白天进入昏暗的环境时,以及在夕阳西下时,它们很可能比具有稳定光色素的生物体有明显的优势。同样地,在同一视网膜内,纤毛光感受器可能比它们的微绒毛同伴享有足够大的优势,就更高的灵敏度和适应黑暗的能力而言,保留两组光感受器几乎没有什么好处。相反,大部分微绒毛细胞失去了它们的感光特性,而变成了特异的投射神经元,尽管有些保留了为昼夜诱捕和其他“非视觉”目的发出信号的作用。
多类视网膜视蛋白和光感受器.在全基因组复制的“2R”轮之前,似乎只有两种锥视蛋白(SWS和LWS)在两类锥视蛋白中表达。然后,作为“2R”复制的结果,SWS视蛋白四倍复制为SWS1, SWS2, Rh2和Rh1视蛋白,在最初应该是四类短到中波敏感锥光感受器中表达。然而,在低光水平下的高灵敏度压力将有利于延长(至少)其中一种视蛋白的meta II寿命,因此光响应可以集成更长的时间。这似乎导致了Rh1视蛋白122和189位点的突变,通过部分屏蔽类视黄蛋白结合位点,使水无法进入,从而延长了meta II的寿命。同样的压力还导致表达Rh1视蛋白的细胞中其他恢复步骤的减缓,因此GRK、arrestin和PDE6的修改版本出现了。尽管LWS视蛋白同样被认为是四倍的,但只有一个副本存活了下来。
即使到了七鳃鳗从我们自己的谱系中分化出来的阶段(即图1中的#5),“真棒”似乎也不太可能进化出来;因此,尽管北半球七鳃鳗的短光感受器表达Rh1视蛋白,并且相当敏感,但它们表现出许多锥状的特性,尚不确定它们是否能够可靠地发送单个光子撞击的信号。我们在下颌脊椎动物中认识到的那种真正的杆,可能在那次分化之后才得到完善。
视网膜双极细胞的进化。在脊椎动物的侧边视网膜以外的光敏区域(即松果体器官、松果体旁器官和顶叶眼),以及盲鳗眼的视网膜中,纤毛光感受器直接与投射神经元进行突触接触,如上所述,投射神经元很可能是微绒毛光感受器的后代。相比之下,脊椎动物视网膜上的锥状和杆状感光细胞通过两极细胞与投射神经元进行交流。对视网膜发育的检查表明,这些双极细胞的分化比所有其他视网膜神经元都要晚,此外,正在发育的双极细胞插入到一个最初由光感受器与神经节细胞直接接触组成的电路中。
这些发现与视网膜双极细胞进化较晚的观点是一致的,在两层视网膜已经建立(包括与投射神经元相连的纤毛光感受器)之后,这些更新的细胞插入到已经存在的神经通路中。这种新的安排极大地提高了视网膜的计算能力,使光感受器信号汇集,以及不同信号的比较,如中心环绕处理和光谱(颜色)比较。
附属结构(晶状体,角膜,肌肉)。在照相机式眼睛中,使视网膜发挥“传感器”功能的辅助结构包括晶状体、角膜和巩膜、虹膜以及眼内和眼外肌肉。此外,可以说是最重要的是,大脑中需要有一个神经处理中心来解释信息并对其采取行动,以及控制眼睛的运动。虽然很明显,这些结构一定是与视网膜的进化同步进化的,但不幸的是,任何关于它们进化的描述都超出了本文的范围。
下颚脊椎动物:第5点之后(500年以来)
圆口目分化后,下颌脊椎动物眼睛的基本bauplan没有发生实质性变化。尽管如此,还是出现了许多改进,其中许多都与眼睛的光学有关,特别是在四足动物出现后,空气界面所需的光学。这里不讨论这些更改。
颌骨脊椎动物视网膜中进化出的一个主要变化涉及到scotopic通路。据我们所知,七鳃鳗和盲鳗都没有颌骨脊椎动物进化出的那种“双”视网膜,它们的真视杆能够可靠地检测单个光子的撞击,并有专门的聚焦视觉系统。beplay体育公司相反,七鳃鳗表达杆状Rh1/RhA视蛋白的光感受器保留了各种锥状特征。根据目前的证据,我认为,现代脊椎动物表现出的对光线的终极敏感性,是由视网膜“计数”击中“真杆”的单个光子的能力所实现的,直到我们的祖先与七鳃鳗和盲鳗的祖先分化后才进化出来的。通过未来的研究来验证这一论断是很重要的。
19总结
以下是被假设为有助于脊椎动物视锥和视杆光感受器及其转导级联进化的主要事件的简要摘要:
1)在后生动物进化的很早就出现了g蛋白级联的原始环核苷酸版本。在这个级联中,一个(非光敏的)GPCR激活了一个g蛋白,该蛋白与一个调节环核苷酸水平的效应酶偶联,从而调节cngc的打开。
2)一类GPCR进化出能以非共价结合视黄醛作为其配体,从而有效地成为视黄醛受体。这发生在平栖动物分化之前。随后,赖氨酸296残基进化,通过席夫碱键与类视黄酮形成共价结合。一个反离子位点(例如E181)也出现了,使希夫碱键被质子化,并产生原始视蛋白,该视蛋白在光谱短波端附近有峰值吸收。
3)表达第一个视蛋白的细胞因此成为原始的后生动物感光细胞。它使用的g蛋白级联与现代纤毛光感受器大致相当,特别是它使用一个环核苷酸作为细胞质信使来调节环核苷酸门控离子通道的打开。
4)这种光感受器细胞进化出来的生物体非常小,还没有可以被称为神经系统的东西。细胞通过局部释放一种化学信使向其他细胞发出信号,可能是褪黑激素和谷氨酸。
5)这个细胞可能不需要11-的来源独联体视网膜,因为视蛋白载脂蛋白可以结合所有-反式视网膜,因为由此产生的视觉色素可以经历光逆转,从活跃的后视视紫红质状态到基态视紫红质(大多数视蛋白都可以,脊椎动物锥视蛋白和杆状视蛋白除外)。
6)通过多次复制,该细胞出现了变体,导致表达c -视蛋白、r -视蛋白或rgr -视蛋白的不同细胞类型。
7)在表达原始视蛋白的细胞中,视蛋白转移到纤毛的质膜上。随着时间的推移,纤毛膜大规模扩张的机制进化了。起初,这些扩张组织得很糟糕;通常它们倾向于平行于纤毛轴(纵向)。
8)在后口动物/脊索动物谱系中,睫状体膜的扩张逐渐演变成更大的组织程度,在头脊索动物分化时,睫状体膜变得扁平(片层状),在腹膜分化时,睫状体膜形成芽状花瓣,随后在脊椎动物出现时,睫状体膜向外展开,与睫状体轴成直角。
9)在早期脊索动物进化过程中,可能只有一种c -视蛋白,很可能属于Opn3家族,在许多方面与r -视蛋白没有太大区别。但在早期脊椎动物进化过程中,出现了这种c视蛋白的许多变体,其中残留在现存脊椎动物中的成员包括视蛋白parietopsin,副视蛋白parinopsin, VA视蛋白和视蛋白pinopsin。
10)在这个过程中,席夫碱的反离子从181号位迁移到113号位,这种重新定位至少在三个方面起了重要作用。首先,它允许释放所有-反式视网膜和因此在黑暗中视觉色素的快速再生(从储存11-独联体视网膜);其次,通过缩短活性元II的使用寿命,提高了锥的时间分辨率;第三,它为g蛋白激活的更高效率铺平了道路,通过进一步的分子内重排,导致螺旋6在meta II状态的巨大倾斜。最后的改进防止了meta II的光逆转,并使all-的释放反式视网膜至关重要。
11)因此,祖先视锥光感受器利用了一种视蛋白(祖先视锥视蛋白),这种视蛋白表达在从纤毛向外侧辐射的板层囊膜中。它需要11-的来源独联体类视黄酮(与现代视锥细胞相似)可能是由Müller细胞提供的。
12)参与级联激活的主要蛋白质已经进化成:(我)祖先转导素,其Gt α亚基从早期Gi复制和修饰,并具有无处不在的β和独特的γ亚基;(2)原始的PDE6环GMP磷酸二酯酶,包括一对相同的催化亚基和一对调节(γ)亚基;和(3)一种四聚体环核苷酸门控通道,由两类亚基组成,α和β。
13)参与反应恢复和调节的主要蛋白已经进化成原始的“锥”成分:GRK7、ARR3、RGS9、GUCY2D、GUCA1C、SLC24A1、RCVRN。
14)第一个锥视蛋白的复制产生了一对视网膜锥视蛋白:一个带有标准E181残基的SWS视蛋白和一个带有H181/K184组合的LWS视蛋白,后者提供了氯离子结合位点和显著的红移吸收。这两种视蛋白在独立但非常相似的视锥光感受器中表达。它们的输出提供了可以用来进行二色处理的信号。
15)附近还存在由共同的祖先光感受器演化而来的微绒毛光感受器。纤毛光感受器的旁分泌释放机制演化为与邻近微绒毛光感受器的突触接触。在某一阶段,纤毛光感受器的性能(传输到微绒毛来源的细胞)变得足以信号所有的照明特征,对生物体是重要的,此后,大多数微绒毛光感受器摆脱了视蛋白和光转导级联,特化为投影神经元。
16)以上所有事件都发生在脊椎动物谱系基础的“2R”两轮全基因组复制之前。
17)由于两个基因组的复制,SWS视蛋白的四倍复制产生了脊椎动物的SWS1、SWS2、Rh2和Rh1视蛋白类。只有一个LWS四倍复制体存活了下来。这五种视蛋白分别在五类视锥细胞中表达。
18)基因组的复制还导致了光转导级联的许多成分的异构体的锥与杆的差异。
19)在表达Rh1视蛋白的锥体光感受器中,许多变化的汇合导致了反应关闭的减缓,这引起了一次光异构化更大的反应。其中一个变化是由于位点122和189的突变导致meta II的寿命增加。这也降低了视紫红质的再生速率,这可能解释了视紫红质的抗羟胺性。
20)细胞外段超微结构的改变,包括质膜内片层的包围,使信使分子的纵向传播更加广泛。
21)在适当的时候,这些变化结合在一起产生了一种光感受器,它能够可靠地向单个光异构化发出信号,因此它值得被称为“第一真棒”。这一阶段是在圆口分化之前还是之后还不确定,但似乎是在圆口分化之后。
22)大约在脊索c视蛋白失去光逆转能力的时候,一种或多种机制出现了在黑暗中类视黄酮的再异构化。这些机制中的第一个可能是现代视锥细胞能够进入的视网膜内部循环。另一种循环,涉及类维生素a在RPE之间的运输,在被囊动物分化时还没有出现,因此(在现存物种中)是一种纯粹的脊椎动物现象。
23)祖先脊索动物前神经管的光敏区向背侧和外侧扩张。睫状体光感受器和投射神经元彼此紧密相连,“伸展”的是投射神经元的轴突,它负责将信息传递到大脑的其他区域。
24)外侧囊泡呈一侧性,向外的一侧只生成神经视网膜的细胞,而近侧只生成提供支持、营养和类视黄素处理功能的上皮细胞,以及吸收多余光线的色素。在进化的那个阶段,侧视网膜与现代盲鳗的视网膜大致相似。
25)作为基因组复制的结果,两种新的视网膜细胞出现了。这些水平细胞和无分泌细胞分别与纤毛光感受器和神经节细胞形成突触连接,其方式介导细胞之间的横向相互作用,并可能介导简单空间对比的提取。
26)同样是基因组复制的结果,另一类新的细胞,视网膜双极细胞,进化了。它在许多方面与睫状体光感受器细胞非常相似,但它没有形成一个外段,而是表达谷氨酸感受器。这个新细胞被纳入了从睫状体光感受器到神经节细胞的神经通路中,从而使视网膜的计算能力大大提高。
27)在视网膜细胞分化和视网膜细胞早期连通性的各种特征中,保留着视网膜细胞进化和视网膜布线进化的遗迹。
(28)脊索动物的空间视觉是逐渐产生的,这不仅是通过光感受器和视网膜的平行变化,还通过成像元素(晶状体)和其他辅助结构的形成,如角膜/巩膜、虹膜、眼内和眼外肌肉,以及通过可以利用空间信息的大脑结构的完善。
(29)空间视觉的出现为生物的生存提供了巨大的价值,但这一过程是缓慢的,经过无数个步骤,从早期脊索动物的一个简单的眼点过渡到脊椎动物式的照相机眼可能需要长达1亿年的时间。
我应该强调的是,上面的简要总结代表了我对脊椎动物光感受器和视网膜进化事件的最佳猜测,这是从文献中大量的实验数据中提炼出来的。对5亿多年前发生的事件提出一个明确的建议的目的是为了刺激进一步的实验,以便检验并从而改进或拒绝特定的建议。我期待着根据未来实验的结果,更新我在上面提出的想法。
确认
我非常感谢Helga Kolb激励我承担这个项目,感谢Nansi Colley, Tom Cronin, John Dowling, Martin Heck, Dan Larhammar, Robert Marc, Ralph Nelson和Yoshinori Shichida对本文草稿的有益评论,感谢Shaun Collin对图片的使用。澳大利亚研究委员会资助项目CE0561903和DP110103294。
1.提出警告,T。生物学中没有任何东西是有意义的,除非从进化的角度来看。美国生物教师,1973年。35: 125 - 129页。[PBS.org]
2.达尔文,C。论自然选择的物种起源.1859年,伦敦:J.默里。502页。[谷歌图书]
3.兰姆,t.d., S.P.科林,还有小E.N.普,脊椎动物眼睛的进化:视蛋白,光感受器,视网膜和眼杯。神经科学,2007。8(12): 960 - 76页。[PubMed]
4.羔羊肉,T.D。,脊椎动物视网膜光接收的进化。生物学杂志,2009。364: p。2911 - 24。
5.阿伦特,D.豪森和G.珀施克,眼睛进化的“分工”模式。生物学杂志,2009。364: p。2809 - 17所示。伦敦皇家学会哲学学报b364, 2809-2817。doi: 10.1098 / rstb.2009.0104
6.沃帕伦斯基,P.和Z.科兹米克,眼睛进化:遗传成分的共同使用和独立招募。生物学杂志,2009。364: 2819 - 32页。[PubMed]
7.拉尔汉玛,D., K.诺德斯特龙,和T.A.拉尔森,脊椎动物杆状和锥状光转导基因的进化。生物学杂志,2009。364: 2867 - 80页。[PubMed]
8.Shichida Y.和T. Matsuyama,视蛋白的进化和光转导。生物学杂志,2009。364: 2881 - 95页。[PubMed]
9.草部,t.g.,等人,脊椎动物视维甲酸循环的演化与起源。生物学杂志,2009。364: 2897 - 910页。[PubMed]
10.科林,s.p.,等人,早期脊椎动物感光器的进化。生物学杂志,2009。364: 2925 - 40页。[PubMed]
11.费恩,g.l., R.哈迪,S.B.劳克林,光转导与光感受器的进化。咕咕叫医学杂志,2010年。20.(3): 114 - 24页。[PubMed]
12.波特,m.l.等人,对视蛋白进化有了新的认识。生物学报,2012。279: 3 - 14页。[PubMed]
13.尼尔森,丧眼睛的进化及其功能基础。Vis >, 2013。30.(1 - 2): 5 - 20页。[PubMed]
14.施瓦布,i.r., R.R. Dubielzig和C. schbert,进化见证:眼睛是如何进化的.2012年,纽约:牛津大学出版社。ISBN 978-0-19-536974-8。306便士。
15.巴特勒,A.B。,脊索动物的进化和颅类动物的起源:新脑中的旧脑。阿娜特Rec, 2000年。261(3): 111 - 25页。[PubMed]
16.巴特勒,A.B.和W. Hodos,比较脊椎动物神经解剖学:进化与适应.2005年第2版,霍博肯,新泽西州:威利-跨学科。715便士。
17.Salvini-Plawen L.和E. Mayer,光感受器和眼睛的进化。另一个星球杂志,1977年。10: 207 - 263页。
18.雅各,F。,进化和修补。科学,1977。196: p。1161 - 6。[Pubmed]
19.戈德史密斯、T.H。视觉光接收的进化修补。Vis >, 2013。30.(1 - 2): 21-37页。[PubMed]
20.Erwin, d.h.,等人,寒武纪难题:动物早期历史中的早期分化和后来的生态成功。科学,2011。334: p。1091 - 7。[PubMed]
21.陈,J.-Y动物王国早期历史的演化情景:中国前寒武纪(埃迪卡拉纪)翁安和早寒武纪茅天山生物群的证据。,在地球与生命,施普林格J.A. Talent,编辑,2012,施普林格。doi: 10.1007 / 978 - 90 - 481 - 3428 - 1 - _10
22.陆地,M.F.和d - e。尼尔森,动物的眼睛.牛津动物生物学系列。2002年,纽约:牛津大学出版社。
23.帕特森,j.r.,等人,寒武纪大型掠食者异鱼目的敏锐视力与复眼的起源。自然,2011年。480: 237 - 40页。[PubMed]
24.普罗尼克,r.e., S.Q.多恩博斯和J.陈,寒武纪辐射的信息景观与感觉生态。古生物学,2010年。36: 303 - 317页。[古生物学]
25.Terakita、。视蛋白。基因组医学杂志,2005年。6(3): p。213。[PubMed]
26.华盛顿的Plachetzki B.M. Degnan和T.H. Oakley,动物视蛋白进化过程中新蛋白相互作用的起源。《公共科学图书馆•综合》,2007年版。2(10): p . e1054。[PubMed]
27.华盛顿的普拉切茨基,C.R.方,还有T.H.奥克利,从祖先环核苷酸门控途径光转导的演化。生物学报,2010。277: 1963 - 9页。[PubMed]
28.Kozmik, Z.等人,由脊椎动物组成的刺胞照相机型眼的组装。美国科学院学报,2008。105(26): 8989 - 93页。[PubMed]
29.Suga, H., V. Schmid和W.J. Gehring,水母视蛋白的进化与功能多样性。咕咕叫医学杂志,2008年。18(1): p。那种。[PubMed]
30.封建,R.等人,后生动物视蛋白进化揭示了动物视觉的一条简单途径。中国科学院学报,2012。109(46): 18868 - 72页。[PubMed]
31.梅森,B.等人,造礁珊瑚多重光转导途径的证据。《公共科学图书馆•综合》,2012年版。7(12): p . e50371。[PubMed]
32.Krishnan, A.等人,GPCRs的起源:哺乳动物如真菌中的视紫红质、粘连、谷氨酸和卷曲GPCRs的鉴定。《公共科学图书馆•综合》,2012年版。7(1): p . e29817。[PubMed]
33.R.M Eakin,感光细胞的进化。冷泉港Symp定量生物学,1965年。30.: 363 - 70页。
34.R.M Eakin,无脊椎动物光感受器的形态。方法Enzymol, 1982。81: 17-25页。[PubMed]
35.范弗雷特伦,j。r。a。库曼斯,光感受器的进化与系统发育。1976年。14: 157 - 168页。[威利]
36.Vanfleteren,黄。单系进化?纤毛诱导光感受器膜。,在视觉细胞的进化, J.A.韦斯特福尔,编辑。1982年,瑞文出版社:纽约。p . 107 - 136。
37.鲁伊斯,M.S.和r。阿纳东,文昌鱼横纹肌光感受器精细结构的若干思考。J Hirnforsch, 1991。32(2): 159 - 64页。[PubMed]
38.布兰登伯格,j.l., R.M.伍拉科特和R.M.埃金,龙眼幼虫眼斑的精细结构(半龙眼门)。Z Zellforsch Mikrosk Anat, 1973。142(1): 89 - 102页。[PubMed]
39.布卢姆,M。pespeselecani(软体动物科,胃足类)个体发生过程中眼睛的改变。动物形态学,1996年。116: 123 - 131页。116年动物形态学,123 - 131。doi: 10.1007 / bf02526944
40.准备好,D.F.和U. Tepass,视网膜形态发生与疾病中的屑依赖性上皮组织,卷10,在人类生物学的最新进展D.S.威廉姆斯,编辑,2004,世界科学出版:河流边缘,新泽西州。7-27页。
41.Lacalli t.c.,文昌鱼幼虫的眼前回路、吻侧感觉通路和大脑组织:来自3D重建的证据。伦敦生物学杂志,1996。351: 243 - 263页。[英国皇家学会]
42.Lacalli t.c.,文昌鱼的感觉系统:远古脊索动物状况的一个窗口。大脑行为研究,2004。64(3): 148 - 62页。[PubMed]
43.戈麦斯,M.P, j·m·安圭拉,e·纳西,文昌鱼黑视素受体的光转导。美国科学院学报,2009。106(22): 9081 - 6页。[PubMed]
44.Koyanagi, M.等人,头脊索黑视素:无脊椎动物视觉细胞和脊椎动物光敏视网膜神经节细胞之间的进化联系。咕咕叫医学杂志,2005年。15(11): 1065 - 9页。[PubMed]
45.安圭拉,j.m.等人,表达黑素的文昌鱼光感受器通过磷脂酶C信号级联转导光。《公共科学图书馆•综合》,2012年版。7(1): p . e29813。PLoS One 7, e29813。doi: 10.1371 / journal.pone.0029813
46.Ferrer, C.等人,解剖文昌鱼微绒毛光感受器的光敏感性决定因素:黑视蛋白信号转导的可能进化意义。J >, 2012。32(50): 17977 - 87页。[PubMed]
47.Vopalensky, P.等人,文昌鱼额眼的分子分析揭示了脊椎动物眼睛的视网膜和色素细胞的进化起源。中国科学院学报,2012。109(38): 15383 - 8页。[PubMed]
48.Sorrentino, M。海鞘幼虫大脑囊泡及其感觉器官的演化。Acta Zool-Stockholm, 2000年。81(3): p。243。
49.巴恩斯,S.N星纹小蝌蚪幼虫光感受器和脑神经节的精细结构(尾索动物亚门;类:海鞘纲)。Z Zellforsch Mikrosk Anat, 1971。117(1): 1 - 16页。[PubMed]
50.埃金,R.M.和A.库达,海鞘蝌蚪感觉受体的超微结构。Z Zellforsch Mikrosk Anat, 1971。112(3): 287 - 312页。[PubMed]
51.戈尔曼,a.l., j。s。麦克里诺兹和s。n。巴恩斯,原始脊索动物的光感受器:精细结构、超极化感受器电位和进化。科学,1971。172: p。1052 - 4。[PubMed]
52.海姆伯格,a.m.,等人,microRNAs揭示了盲鳗、七鳃鳗和颌口目动物的相互关系以及祖先脊椎动物的性质。美国国家科学院学报,2010。107(45): 19379 - 83页。[PubMed]
53.Zintzen, V.等人,盲鳗的捕食行为和粘液防御机制。2011年Sci代表。1: p。131。[PubMed]
54.D.R Newth,地粘藤对光反应的研究。实验生物学杂志,1955年。32(1): 4页。
55.小林,H。盲鳗眼睛的感光功能。国立水产大学学报(前身为马关水产大学学报),1964年。13: 67 - 83页。http://www.fish-u.ac.jp/kenkyu/sangakukou/kenkyuhoukoku/13/13-2-9.pdf
56.吊坠,N.A.和J.M.乔根森,盲鳗的生物学,在盲鳗的生物学J.M. Jørgensen等人,编辑。1998,查普曼&霍尔:伦敦;纽约。p . 541 - 546。
57.开始,K。盲鳗视网膜:比较太平洋盲鳗(Polistotrema stouti)和大西洋盲鳗(Myxine glutinosa)受体和上皮细胞的电子显微镜研究。Z Zellforsch Mikrosk Anat, 1971。121(2): 249 - 69页。[PubMed]
58.开始,K。圆口纲脊椎动物视网膜。《感觉生理学手册》,1977。7(5): 47 - 66页。
59.芬霍姆,B.和K.霍姆伯格,三种盲鳗属(Eptatretus, Paramyxine和Myxine)的眼睛——退行性进化的一个例子。视觉Res, 1975。15(2): 253 - 9页。[PubMed]
60.布洛克,t.h., j·k·摩尔和r·d·菲尔兹,髓鞘的进化:七鳃鳗和盲鳗都缺乏髓鞘。>, 1984。48(2): 145 - 8页。[PubMed]
61.R·舒尔茨、E.C.伯科维茨和D.C.·皮斯,七鳃鳗脊髓的电镜图。J Morphol, 1956。98(2): 251 - 273页。
62.Kusunoki, T. F. Amemiya,盲鳗的视网膜投影。大脑Res, 1983。262(2): 295 - 8页。[PubMed]
63.威希特,h·和r·g·诺卡特,太平洋盲鳗的视网膜移和视网膜顶投影。大脑行为进化,1990。36(5): 315 - 28页。[PubMed]
64.哈特,S.等人,小鼠视网膜神经节细胞表达黑视素的中央投射。神经科学杂志,2006。497(3): 326 - 49页。[PubMed]
65.艾伦,B.M。,stouti的眼睛。阿娜特。澳新银行,1905年。26: 208 - 211页。在http://www.biodiversitylibrary.org/ia/anatomischeranze26anat
66.普,G.A.和J.E.道林,七鳃鳗幼虫松果体光感受器细胞的解剖和生理特征。J Neurophysiol, 1981。46(5): 1018 - 38页。[PubMed]
67.R.M Eakin,第三眼.1973年,伯克利,加利福尼亚大学出版社。157页。谷歌图书]
68.库斯米克,C., P.马尔基亚法瓦和E.斯特雷特托伊,鳟鱼松果体光感受器的光响应和光适应。生物学报,1992。248: 149 - 157页。[英国皇家学会]
69.森田,M. Tabata和S. Tamotsu,松果体光感受器和神经节细胞的细胞内反应和输入电阻变化。神经科学研究,1985。2: s79 - 88页。[PubMed]
70.兰姆,T.D.和e。n。普,Jr,两栖动物光感受器光转导激活步骤的定量描述。杂志,1992年。449: 719 - 58页。[PubMed]
71.Koyanagi, M.等人,七鳃鳗松果体中的双稳态紫外色素。美国国家科学院,2004。101(17): 6687 - 91页。[PubMed]
72.内田,K.和Y.森田,光感松果体器官中紫外线敏感细胞的细胞内反应。大脑Res, 1990。534(1 - 2): 237 - 42页。[PubMed]
73.Burkhardt,地方检察官。锥状光感受器的突触反馈、去极化和颜色对抗。Vis >, 1993。10(6): 981 - 9页。[PubMed]
74.Meissl H. T. Nakamura和G. Thiele,金鱼光感松果体的神经反应机制。生物化学物理杂志,1986。84(3): 467 - 73页。[PubMed]
75.弗里茨奇,B.和S.P.科林,银七鳃鳗视网膜中两群神经节细胞和视网膜瓣纤维的树突状分布。Vis >, 1990。4(6): 533 - 45页。[PubMed]
76.里的年轻,原始颌口鱼眼外肌的数量和排列:来自已灭绝的皮目鱼类的证据。生物学快报》,2007年版。4: 110 - 114页。[PubMed]
77.年轻的GC(2008)。脊椎动物眼睛的早期进化:化石证据。进化:教育与拓展1,427 -438。doi: 10.1007 / s12052 y - 008 - 0087 - (施普林格]
78.Fritzsch, B。,视觉系统系统发育的个体发生线索beplay体育公司,在视觉系统的变化:中枢神经系beplay体育公司统的成熟和衰老P. Bagnoli和W. Hodos,编辑,1991,全会出版社:纽约。33-49页。[施普林格]
79.Fritzsch b和J.C. Glover,后口中枢神经系统的进化:发育模式过程与细胞规范过程的插入,卷2,在神经系统的进化:一个全面的参考, j·h·卡斯,编辑。2007,爱思唯尔学术出版社:阿姆斯特丹;波士顿。
80.Nivison-Smith, L.等,南半球七鳃鳗视网膜氨基酸神经化学。《公共科学图书馆•综合》,2013年版。8(3): p . e58406。[PubMed]
81.科林,s.p.,等人,古代色觉:远古脊椎动物的多重视蛋白基因。咕咕叫医学杂志,2003年。13(22): R864-5页。[PubMed]
82.Pisani, D.等人,在脊椎动物的最后一个共同祖先身上有微光视觉的分子证据。咕咕叫医学杂志,2006年。16(9): p。R318-9;作者R320回复。[PubMed]
83.科林,s.p.,等人,南半球七鳃鳗视网膜光感受器的形态和光谱吸收特性。Vis >, 2003。20.(2): 119 - 30页。[PubMed]
84.迪克森,D.H.和T.R.科拉德,七鳃鳗视网膜的发育:变质前的隐卵眼。阿纳特,1979年。154(3): 321 - 36页。[PubMed]
85.科林,S.P.和A.E.O.特雷兹,脊椎动物颜色辨别的进化及其对视觉交流的意义,在沟通在鱼, F. Ladich,编辑。2006,科学出版社:恩菲尔德,NH。p . 303 - 335。
86.穆拉多夫等人,七鳃鳗(lamprey Petromyzon marinus)的PDE6:对脊椎动物视觉效应进化的启示。生物化学,2007年。46(35): 9992 - 10000页。[PubMed]
87.穆拉多夫等人,七鳃鳗长、短光感受器中表达独特的转导因子。视觉Res, 2008。48(21): 2302 - 8页。[PubMed]
88.史密斯,j.j.等人,海洋七鳃鳗(Petromyzon marinus)的基因组测序为脊椎动物进化提供了深刻的见解。Nat麝猫,2013年。45(4): p. 415- 21,421e1 -2。[PubMed]
89.戈瓦尔多夫斯基,V.和D.利恰科夫,七鳃鳗的视觉细胞和视觉色素。[A], 2004。154(2): 279 - 286页。[施普林格]
90.戴维斯,w.l.等人,溯河七鳃鳗视觉色素基因表达的功能表征、调节与调控。美国实验生物学学会联合会J, 2007。21(11): 2713 - 24页。[PubMed]
91.Ebrey, T.和Y. Koutalos,脊椎动物的感光细胞。视网膜眼科研究,2001。20.(1): 49 - 94页。
92.Imai, H.,等人,从纯化的鸡视锥色素到小鼠视紫红质的视锥色素的分子性质。光化学与生物学报,2005。4(9): 667 - 74页。[PubMed]
93.里特,E.等人,视紫红质向活性后视紫红质II状态的转变打开了一个与希夫碱异构化相关的新的光诱导通路。生物化学杂志,2004。279(46): 48102 - 11页。[PubMed]
94.戴维斯,w·l, m·w·汉金斯,r·g·福斯特,脊椎动物古代视蛋白和黑视蛋白:发散辐照度探测器。光化学与生物学报,2010。9(11): 1444 - 57页。[PubMed]
95.Koyanagi, M.等人,脊椎动物Opn3的同源物可能在非光敏感组织中充当光传感器。美国科学院学报,2013。110(13): 4998 - 5003页。[PubMed]
96.酒井,K,等人,壁菌素的光化学性质。生物化学,2012年。51(9): 1933 - 41页。[PubMed]
97.Terakita, A.等人,视紫红质家族分子进化中的反离子置换。细胞生物学杂志,2004。11(3): 284 - 289页。[PubMed]
98.佐藤,K.等,脊椎动物视蛋白具有介于脊椎动物和无脊椎动物视觉色素之间的分子性质。生物化学,2011年。50(48): 10484 - 90页。[PubMed]
99.冈野,T,吉泽T,深田y,Pinopsin是一种鸡松果体感光分子。自然,1994年。372: p。94 - 7。[PubMed]
One hundred.Nakamura, A.等人,视视素在杆状和锥状视觉色素间的嵌合性质。生物化学,1999年。38(45): 14738 - 45页。[PubMed]
101.Tsutsui, K.和Y.七田,希夫碱反离子在视紫质中的多重功能。光化学与生物学报,2010。9(11): 1426 - 34页。[PubMed]
102.冢本,H.和A. Terakita双稳定颜料的多样性与功能特性。光化学与生物学报,2010。9(11): 1435 - 43页。[PubMed]
103.Terakita, A., E. Kawano‐Yamashita和M. Koyanagi,视蛋白的进化与多样性。膜传输与信号转导,2012。1(1): 104 - 111页。
104.陈,m.h.,等人,视网膜视色素在光激活后从锥状视色素中迅速释放生物化学,2012年。51(20): 4117 - 25页。[PubMed]
105.冢本,H.等人,不同视紫质在光诱导下构象变化的程度与其激活G蛋白的能力有关。生物化学杂志,2009。284(31): 20676 - 83页。[PubMed]
106.Choe, h.w.,等人,后视紫红质晶体结构2。自然,2011年。471(7340): 651 - 5页。[PubMed]
107.Scheerer, P.等人,视蛋白与g蛋白相互作用构象的晶体结构。自然,2008年。455: 497 - 502页。[PubMed]
108.霍夫曼,k.p.等人,G蛋白偶联受体在工作:视紫红质模型。生物化学,2009。34(11): 540 - 52页。[PubMed]
109.Yan, E.C,等,G蛋白偶联受体视紫红质的光激活中的视网膜反离子开关。美国科学院学报,2003。One hundred.(16): 9262 - 7页。[PubMed]
110.Piechnick, R.等人,通道突变对视蛋白中视网膜配体摄取和释放的影响。中国科学院学报,2012。109(14): 5247 - 52页。[PubMed]
111.Ohno、S。基因复制进化.1970年,柏林,纽约,斯普林格-弗拉格。十五、160 p。
112.埃克尔,j.l.,等人,表达黑素的视网膜神经节细胞光感受器:细胞多样性及其在模式视觉中的作用。神经元,2010年。67(1): 49-60页。[PubMed]
113.施密特,t.m.,陈世凯和S.哈特,本光敏性视网膜神经节细胞亚型多,功能多样。趋势>,2011。34(11): 572 - 80页。[PubMed]
114.Bowmaker, j.k.。脊椎动物视觉色素的进化。视觉Res, 2008。48(20): 2022 - 41页。[PubMed]
115.Okano, T.,等人,鸡视锥视觉色素的基本结构:脊椎动物视紫红质是由视锥视觉色素进化而来的。美国科学院学报,1992。89(13): 5932 - 6页。[PubMed]
116.Yamashita, T.,等,l族锥视色素的氯离子依赖性光谱调谐机制。生物化学,2013年。52(7): 1192 - 7页。[PubMed]
117.王,Z, A.B. Asenjo和D.D. Oprian,人类红色和绿色视色素中Cl(-)结合位点的鉴定。生物化学,1993年。32(9): 2125 - 30页。[PubMed]
118.戴维斯,W.I,等人,中长波长敏感视色素的阴离子敏感性及光谱调谐。细胞与生命科学,2012。69(14): 2455 - 64页。[PubMed]
119.戴维斯,w.l.等人,鸭嘴兽的视觉色素:哺乳动物色彩视觉的新途径。咕咕叫医学杂志,2007年。17(5): R161-3页。[PubMed]
120.戴维斯,w。i。科林,d。m。亨特,颅类视光色素的分子生态学及其适应性。2012年摩尔生态。21(13): 3121 - 58页。[PubMed]
121.诺德斯特龙,K, T.A.拉尔森和D.拉汉玛,早期脊椎动物进化过程中光转导基因的大量复制与染色体块(block)复制相关。基因组学,2004年。83(5): 852 - 72页。[PubMed]
122.Golobokova, E.Y.和V.I. Govardovskii,视觉色素原位光解后期:视锥细胞vs视杆细胞。视觉Res, 2006。46(14): 2287 - 97页。[PubMed]
123.川村,横山s和s,美洲变色龙视觉和非视觉色素的功能表征。视觉Res, 1998。38(1): 37-44页。[PubMed]
124.比克曼,C.等人,基础哺乳动物针鼹(Tachyglossus aculeatus)杆视色素的功能特征。Vis >, 2012。29(4 - 5): 211 - 7页。[PubMed]
125.亨特,d.m.等人,深海鱼类杆状视觉色素光谱调谐的分子基础。实验生物学杂志,2001。204(Pt 19):第3333-44页。[PubMed]
126.丘坤威和R.C.哈迪,光转导的主题和变化。细胞,2009。139(2): 246 - 64页。[PubMed]
127.哈迪,r·c·k·弗兰兹,果蝇光感受器的感光反应。科学,2012。338: p。260 - 3。[PubMed]
128.大川,H.,等人,哺乳动物光感受器在黑暗和光明下的ATP消耗。咕咕叫医学杂志,2008年。18(24): 1917 - 21页。[PubMed]
129.斯坦伯格,r.h., S.K.费雪,D.H.安德森,脊椎动物光感受器的圆盘形态发生。神经科学杂志,1980。190(3): 501 - 8页。[PubMed]
130.年轻,R.W。,视觉细胞和更新的概念。投资眼科视觉科学,1976。15(9): 700 - 25页。[IOVS]
131.赫克特,S。S。施莱尔和m。h。皮伦,能量,量子和视觉。物理学杂志,1942年。25(6): 819 - 40页。[PubMed]
132.贝勒,地方检察官,T.D.兰姆,丘家卫,视网膜杆对单光子的反应。杂志,1979年。288: p。613 - 34。[PubMed]
133.羔羊肉,T.D。,光转导:视锥细胞的适应。,卷3,在眼睛百科全书, d.a Dartt, J.C. Besharse, R. Dana,编辑。2010,学术出版社:牛津。p . 354 - 360。
134.川村,S.和S.立banaki,杆状和锥状光感受器:它们生理差异的分子基础。生物化学与物理学报,2008。150(4): 369 - 77页。[PubMed]
135.Tachibanaki, S.等人,鲤鱼球果中转导素的低活化和快速失活。生物化学杂志,2012。287(49): 41186 - 94页。[PubMed]
136.Kenkre, j.s.,等人,在漂白暴露消失时,人体锥体循环电流恢复极快。杂志,2005年。567(Pt 1):第95-112页。[PubMed]
137.阿伦特,D. J.维特布罗德,重建Urbilateria的眼睛。国际生物学杂志,2001。356: 1545 - 1563页。[PubMed]
138.Koyanagi, M.等人,水母的视觉始于opsin-Gs级联介导的cAMP信号。美国科学院学报,2008。105(40): 15576 - 15580页。[PubMed]
139.兰姆,T.D.和e。n。普,Jr,光转导,黑暗适应,和视紫红质再生,proctor讲座。投资眼科视觉科学,2006。47(12): 5137 - 52页。[IOVS]
140.T.G Wensel,光感受器外节的信号转导膜复合体。视觉研究,2008年。48(20): 2052 - 2061页。[PubMed]
141.古列维奇,V.V.等人,光感受器细胞中视觉阻滞剂的功能周期。视网膜眼科研究,2011。30.(6): 405 - 430页。[PubMed]
142.舒尔茨,h.l.,等人,视网膜组-定义成年哺乳动物视网膜/视网膜色素上皮的参考转录组。BMC基因组学,2004年。5(1): 50页。[PubMed]
143.Vogalis, F.等人,锥体异位表达量具体的克量蛋白质量斑马鱼体内的偶联受体激酶GRK7导致光敏性降低和反应改变。杂志,2011年。589(9): 2321 - 2348页。[PubMed]
144.J.I Korenbrot,杆状和锥状光感受器光响应的速度、灵敏度和稳定性:事实和模型。视网膜眼科研究,2012。31(5): 442 - 466页。[PubMed]
145.穆拉多夫,H·k·博伊德,N.O.阿特米耶夫,棒状磷酸二酯酶-6 PDE6A和PDE6B亚基在酶上是等效的。生物化学杂志,2010。285(51): 39828 - 34页。[PubMed]
146.久臣,德永,脊椎动物光转导蛋白的分子进化。比较生物化学和生理学B部分:生物化学和分子生物学,2002。133(4): 509 - 522页。[PubMed]
147.Zhang Z.和N.O. Artemyev,磷酸二酯酶6 γ亚基抑制的决定因素。生物化学,2010年。49(18): 3862 - 3867页。[PubMed]
148.彭,C。d。里奇,和瓦纳姆医生,异聚体锥环核苷酸门控通道的亚基构型。神经元,2004年。42(3): 401 - 410页。[PubMed]
149.穆舍吉安,A·v·古列维奇,和e·v·古列维奇,G蛋白偶联受体激酶的起源和进化。《公共科学图书馆•综合》,2012年版。7(3): p . e33806。[PubMed]
150.古列维奇,E.V.和V.V.古列维奇,抑制素:无处不在的细胞信号通路的调节因子。基因组生物学,2006。7(9): 236页。[PubMed]
151.阿尔瓦雷斯,公元关于逮捕素和视紫红质的起源。BMC进化生物学,2008。8(1): 222页。[PubMed]
152.Kawano-Yamashita, E.等,β -止动素对七鳃鳗松果体中非漂白色素副菠萝素的功能调节作用。《公共科学图书馆•综合》,2011年版。6(1): p . e16402。[PubMed]
153.Lagman, D.等人,早期脊椎动物四倍化中转导亚基基因家族的扩展。基因组学,2012年。[PubMed]
154.泰勒,C.W.和R.D.哈默,视觉调制灵敏度分析。四、Ferry-Porter定律的有效性。JOSA, 1990。7(4): 743 - 758页。[PubMed]
155.诺曼,R.A.和F.S.维尔布林,视网膜敏感性的控制。脊椎动物视杆和视锥的光和暗适应性。Gen Physiol, 1974。63(1): 37 - 61页。[PubMed]
156.Burkhardt,地方检察官。龟视网膜原位锥光感受器的光适应和光色素漂白。J >, 1994。14(3 Pt 1):第1091-105页。[PubMed]
157.t.i. Rebrik等人,CNG-modulin:一种新的钙依赖的锥光感受器cgmp门控离子通道配体敏感性调节剂。J >, 2012。32(9): 3142 - 53页。[PubMed]
158.Mahroo o和T. Lamb,漂白曝光后人体视网膜电图的恢复:色素再生动力学的估计。杂志,2004年。554(2): 417 - 437页。[PubMed]
159.托马斯·m·t·兰姆,视网膜电图a波测定人视杆感光器的光适应和暗适应。杂志,1999年。518(2): 479 - 496页。[PubMed]
160.兰姆,t·e·普,黑暗适应和视觉的类视黄酮循环。视网膜眼科研究,2004。23(3): 307 - 380页。[PubMed]
161.范Hateren、H。灵长类视锥细胞和水平细胞的反应动力学和适应的细胞和分子模型。J Vis, 2005。5(4)。PubMed]
162.Van Hateren, J.和T. Lamb,人视锥细胞的光电流响应快且单相。BMC神经科学,2006。7(1): 34页。[PubMed]
163.拉格纳多,L.瑟韦托,P.麦克诺顿,虎蝾螈视网膜杆外节段的钙稳态。杂志,1992年。455(1): 111 - 142页。[PubMed]
164.斯泰尔斯,W。眼睛光谱灵敏度曲线的物理解释,在敬拜眼镜制造商公司光学大会会刊.1948年,伦敦奇观制造公司。p . 97 - 107。
165.刘易斯,P。长波光谱灵敏度曲线的理论解释。杂志,1955年。130(1): 45页。[PubMed]
166.巴洛,H。浦肯野移位和视网膜噪声。自然,1957年。179: 255 - 256页。[PubMed]
167.Ala-Laurila, P.等人,视觉颜料的光活化能与吸收光谱的关系。视觉Res, 2004。44(18): 2153 - 2158页。[PubMed]
168.罗,D.-G。、等。光和热激活视觉色素。科学,2011。332: 1307 - 3012页。[PubMed]
169.Ala-Laurila P. K. Donner和A. Koskelainen视觉色素的热激活和光激活。2004年Biophys J。86(6): 3653 - 3662页。[PubMed]
170.Gozem, S.等人,棒状光感受器热噪声的分子机制。科学,2012。337: 1225 - 1228页。[PubMed]
171.Ma, J.,等。视色素在视杆和视锥感光器中都有表达的视觉色素神经元,2001年。32(3): 451 - 61页。[PubMed]
172.马修斯,G。,蛙蛙蝌蚪单视网膜杆的光电流。视觉Res, 1984。24(8): 821 - 5页。[PubMed]
173.兰姆,T.D.和E.J.西蒙,龟锥的电噪声分析。杂志,1977年。272(2): 435 - 68页。[PubMed]
174.兰姆,T.D.和E.J.西蒙,龟光感受器细胞间偶联与电噪声的关系。杂志,1976年。263(2): 257 - 86页。[PubMed]
175.Schneeweis, D.M.和J.L. Schnapf,猕猴锥光感受器的光电压:适应性、噪声和动力学。J >, 1999。19(4): 1203 - 16页。[PubMed]
176.贝勒,地方检察官,g·马修斯,丘家卫,蟾蜍视网膜棒外节电暗噪声的两个组成部分。杂志,1980年。309: 591 - 621页。[医学图书馆]
177.惠特洛克,G.G.和T.D.兰姆,蟾蜍杆单光子响应时间过程的变异性:视紫红质的活性终止。神经元,1999年。23(2): 337 - 51页。[PubMed]
178.哈默,r.d.,等人,建立脊椎动物光导杆的统一模型。Vis >, 2005。22(4): 417 - 36页。[PubMed]
179.哈默,r.d.,等人,活化的视紫红质磷酸化的多个步骤可以解释脊椎动物杆状单光子反应的可重复性。普通物理杂志,2003。122(4): 419 - 44页。[PubMed]
180.格罗斯,op,小e。n。普,还有法医伯恩斯,钙对cGMP合成的反馈强烈减弱了由长视紫红质寿命驱动的单光子响应。神经元,2012年。76(2): 370 - 82页。[PubMed]
181.坎吉亚诺,L.等人,小鼠视网膜中视杆和视锥细胞的光电压。杂志,2012年。590(Pt 16):第3841-55页。[PubMed]
182.兰姆,t.d., P.A.麦克诺顿,丘家威,蟾蜍杆外节激活与背景脱敏的空间分布。杂志,1981年。319: 463 - 96页。[PubMed]
183.墙壁,G.L。,爬行动物的视网膜。点。1934年j .角膜切削。17: 892 - 915页。
184.墙壁,G.L。,脊椎动物的眼睛和它的适应性辐射.克兰布鲁克科学研究所。1942年,布卢姆菲尔德山,密歇根:克兰布鲁克科学研究所。785页。
185.克莱因施密特,j和j。e。道林,壁虎光感受器在适应光和暗过程中的细胞内记录。医学杂志,1975。66(5): 617 - 48页。[PubMed]
186.里斯波利,G, W.A. Sather和P.B. Detwiler,蜥蜴视网膜透析脱离杆外节段的视觉转导。杂志,1993年。465: 513 - 37页。[PubMed]
187.滚,B。,壁虎视觉-视觉细胞,进化和生态约束。J Neurocytol, 2000。29(7): 471 - 84页。[PubMed]
188.吉田,M。夜行壁虎光感受器外节段通透性的一些观察。视觉Res, 1978。18(2): 137 - 43页。[PubMed]
189.小岛,D,等人,锥体视觉色素存在于壁虎杆状细胞中。美国科学院学报,1992。89(15): 6841 - 5页。[PubMed]
190.张x, T.G. Wensel,袁c,Tokay壁虎光感受器通过锥状蛋白质实现杆状生理学。Photochem Photobiol, 2006年。82(6): 1452 - 60页。[PubMed]
191.里普斯,h·j·e·道林,鳐视网膜光感受器的结构特征和适应性。中国农业科学,1990。5: p。46 - 54岁。[PubMed]
192.道林,J.E.和h。里普斯,关于鳐鱼视网膜的双重性。中国农业科学,1990。5: 55 - 65页。[PubMed]
193.道林,J.E.和h。里普斯,鳐鱼光感受器的适应性。医学杂志,1972年。60(6): 698 - 719页。[PubMed]
194.康沃尔,m.c.等人,滑板光感受器的膜电流响应。J Gen Physiol, 1989。94(4): 633 - 47页。[PubMed]
195.王,js。v.j.n Kefalov,视锥特定的视觉周期。视网膜眼科研究,2011。30.(2): 115 - 128页。[PubMed]
196.李鸿源岛,视杆和视锥视觉周期中的维生素A代谢。Annu Rev Nutr, 2012。32: 125 - 45页。[PubMed]
197.凯勒,j.j.等人,视网膜穆勒胶质细胞中DES1作为维生素a异构酶的鉴定。Nat Chem Biol, 2013。9(1): 30-6页。
198.Albalat, R。视觉周期遗传机制的进化:脊椎动物眼睛的新事物?中华分子生物学杂志,2012。29(5): 1461 - 9页。[PubMed]
199.Poliakov, E.等人,脊椎动物视觉周期中视黄酮异构化机制的起源与演化:来自无颚脊椎动物的提示。《公共科学图书馆•综合》,2012年版。7(11): p . e49975。[PubMed]
200.Takimoto, N.等,脊椎动物视觉周期的起源。肠肠Ciona中RPE65和-胡萝卜素15,15 ' -单加氧酶同源物分布明显。Photochem Photobiol, 2006年。82(6): 1468 - 74页。[PubMed]
201.D.G Stavenga,无脊椎动物视觉色素的暗再生,在光感受器光学A.W. Synder和R. Menzel,编辑,1975,施普林格。p . 290 - 295。
202.王,X.,等,果蝇对酶促视觉循环的要求。咕咕叫医学杂志,2010年。20.(2): 93 - 102页。[PubMed]
203.贝尔,K.vEntwicklungsgeschichte der thiere: Beobachtung and reflexition。Borntrager,哥尼斯堡,1828年。1837.{互联网档案馆]
204.Brauckmann、S。卡尔·恩斯特·冯·贝尔(1792-1876)和进化论。国际发展生物学杂志,2012。56(9): 653 - 60页。[PubMed]
205.海克尔,E。Naturliche schopfungsgeschichte。Gemeinverständliche wissenschaftliche vorträge über die entwickelungslehre im allgemeinen und diejenige von Darwin,歌德和拉马克im besonderen.5.动词。aufl。Mit dem porträt des verfassers (nach einer photographie)和Mit 16 tafeln, 19 holzschnitten, 18 stammbäumen和19 systematischen tabellen。1874年,柏林:G.雷默。Xlvi, 2,688 p. [谷歌图书]
206.R.M Eakin,第三只眼睛:一个百年的动物学迷向电子显微镜学家和神经生理学家揭示了它的秘密。1970年美国科学家。58(1): 73 - 79页。
207.Vigh, B.,等。折叠视网膜的松果体器官:视蛋白的免疫细胞化学定位。细胞杂志,1998。90(9): 653 - 9页。[PubMed]
208.A.莱辛巴赫和S.R.罗宾逊,视网膜组织和发育的系统发育限制。视网膜眼科研究,1995。15: 139 - 171页。
209.Vigh, B.,等。大脑深处、松果体器官和视网膜的非视觉光感受器。Histol Histopathol, 2002年。17(2): 555 - 90页。[PubMed]
210.埃克斯特伦,P.和H.梅斯尔,光感松果体器官的进化:神经内分泌光感受器的命运。生物学杂志,2003。358(1438): 1679 - 700页。
211.克莱恩特区脊椎动物松果体的进化:AANAT假说。Chronobiol Int, 2006年。23(1 - 2): 5 - 20页。[Pubmed]
212.Mano, H.和Y. Fukada,中位第三眼:松果体追溯脊椎动物感光器官的进化。Photochem Photobiol, 2007年。83(1): 11 - 8页。[PubMed]
213.埃金,R.M.和J.A.韦斯特福尔蜥蜴顶眼精细结构的进一步观察。生物化学杂志,1960。8: 483 - 99页。[PubMed]
214.马丁内斯-莫拉莱斯,J.R.和j.j Wittbrodt,塑造脊椎动物的眼睛。Curr Opin Genet Dev, 2009。19(5): 511 - 7页。[PubMed]
215.Valleix, S.等人,FOXE3基因纯合子无义突变是导致人类先天性原发性无晶状体的一个原因。Am J Hum Genet, 2006。79(2): 358 - 64页。[PubMed]
216.Eiraku, M.等人,三维培养中的自组织光杯形态发生。自然,2011年。472(7341): 51-6页。[PubMed]
217.Bharti, K.等人,一个涉及PAX6、MITF和WNT信号通路的调控环控制着视网膜色素上皮的发育。公共科学图书馆麝猫,2012年。8(7): p . e1002757。[PubMed]
218.英格兰,s.j.等,前脑的动态命运图显示了脊椎动物的眼睛是如何形成的,并解释了独眼畸形的两个原因。发展,2006年。133(23): 4613 - 7页。[PubMed]
219.Rembold, M.等人,单个细胞的迁移是视神经囊外翻的动力。科学,2006。313: p。1130 - 4。[PubMed]
220.关,k.m.,等人,脊椎动物的眼睛是由复杂的细胞运动形成的。发展,2012年。139(2): 359 - 72页。[PubMed]
221.鲁宾森,K.和H.凯恩,海七鳃鳗幼体视网膜的神经分化。Vis >, 1989。3.(3): 241 - 8页。[PubMed]
222.该,K。七鳃鳗发育中的视觉系统与变态。beplay体育公司一般人,1990。21(7): 1123 - 35页。[PubMed]
223.Meyer-Rochow, v。b。和d。Stewart,七鳃鳗(环口目)幼虫及幼虫后眼超微结构的研究进展,特别以南地鳗(灰)为重点。Microsc Res Tech, 1996。35(6): 431 - 44页。[PubMed]
224.迪克森,d.h.,格雷夫斯检察官,莫伊尔斯先生,发育中的七鳃鳗眼部角膜分裂。阿纳特,1982年。165(1): 83 - 98页。[PubMed]
225.里的价格,黏菌样菌(Bdellostoma stouti Lockington)发育的一些要点。解剖学家,解剖学家,安泽格,1896。6: 81 - 86页。[谷歌图书]
226.院长,B。,关于加利福尼亚盲鳗的发展,大盲鳗,Lockington。显微科学,1897。40: 269 - 280页。http://jcs.biologists.org/content/s2-40/158/269
227.院长,B。,小圆孔虫的胚胎学研究:从卵、节段到孵化的粘液样体发育的概论,在Festschrift zum siebensigsten Geburtstag von Carl von Kupffer.1899年,古斯塔夫·费舍尔:耶拿。[谷歌图书]
228.冯枯氏,C。Zur Kopfentwicklung von Bdellostoma。研究人员zur vergleichenden entwicklungsgeschicte des Kopfes der Kranioten,重4。1900年,慕尼黑:雷曼。www.worldcat.org/title/zur-kopfentwicklung-von-bdellostoma/oclc/250304157
229.Stockard镉比,短吻鳄晶状体的胚胎史与近期实验的关系。阿纳特,1906年。6(1): 511 - 515页。[威利]
230.太田,k.g.,仓乐,仓谷,参考神经嵴进化的盲鳗胚胎学。自然,2007年。446(7136): 672 - 5页。[PubMed]
231.阿伦特、D。眼睛和感光细胞类型的进化。国际发展生物学杂志,2003。47(7 - 8): 563 - 71页。[PubMed]
232.Sernagor, E.等人,视网膜的发展.2006,剑桥:剑桥大学出版社,383页。
233.Agathocleous M.和W.A. Harris,脊椎动物视网膜从祖细胞到分化细胞。细胞发育生物学,2009。25: 45 - 69页。[PubMed]
234.斯瓦洛普,A·金,d·福雷斯特,哺乳动物视网膜光感受器发育和内稳态的转录调控。神经科学,2010。11(8): 563 - 76页。[PubMed]
235.瑞茜,B.E。,视网膜和视神经通路的发育视觉Res, 2011。51(7): 613 - 32页。[PubMed]
236.派波特、D.H。视网膜神经发生,在视网膜的发展, E. Sernagor等人,编辑。2006,剑桥大学出版社:剑桥。30-58页。
237.Rapaport, d.h.等人,大鼠视网膜细胞发生的时间和地形。神经科学杂志,2004。474(2): 304 - 24页。[PubMed]
238.王L.L.和D.H. Rapaport,用克隆分析方法确定黄爪蟾视网膜祖细胞能力。发展,2009年。136(10): 1707 - 15页。[PubMed]
239.戈麦斯,f.l.,等人,体外重建大鼠视网膜祖细胞谱系揭示了细胞命运决定的惊人的随机程度。发展,2011年。138(2): 227 - 35页。[PubMed]
240.他,J.等人,可变的克隆如何建立一个不变的视网膜。神经元,2012年。75(5): 786 - 98页。[PubMed]
241.Godinho, L.等人,非顶端对称分裂是体内发育中的视网膜水平细胞层形成的基础。神经元,2007年。56(4): 597 - 603页。[PubMed]
242.道林,J.E视网膜:大脑中可接近的部分.2012年修订版,剑桥,马萨诸塞州:哈佛大学出版社贝尔纳普出版社。十六、355 p。
243.Manns M.和B. Fritzsch,大脑中的眼睛:视黄酸影响人眼的形态发生和轴突的路径选择,但不影响视网膜的分化。>, 1991。127(2): 150 - 4页。[PubMed]
244.约翰逊,p.t., M.A.雷文,还有B.E.里斯,貂鼠胆碱能无分泌细胞早期消融后内丛状层内瞬态光感受器定位的中断。Vis >, 2001。18(5): 741 - 51页。[Pubmed]
245.约翰逊,p.t.等人,在发育过程中,杆状和锥状突起到内丛状层。神经科学杂志,1999。414(1): 1 - 12页。[Pubmed]
246.Dodt、E。顶眼:低等脊椎动物的顶眼(松果体和顶骨器官,在大脑中的视觉中心(感觉生理学手册)荣格,编辑,1973,施普林格。p . 113 - 140。
247.埃克斯特伦,P.和H.梅斯尔,硬骨鱼松果体器官中经生理鉴定的光感受器细胞和超极化中间神经元的细胞内染色。神经科学,1988。25(3): 1061 - 70页。[PubMed]
248.内田,K., T.中村和Y.森田,七鳃鳗松果体光感受器到发光型神经节细胞的信号传递。神经科学,1992。47(1): 241 - 7页。[PubMed]
249.mef, A。,[针叶鱼大脑细胞结构的电镜研究]。Z Zellforsch Mikrosk Anat, 1973。139(4): 511 - 32页。[PubMed]
250.鲁伊斯,S. R.阿纳东,文昌鱼(鳃口鱼,头索鱼)大脑中层状细胞的精细结构。细胞组织研究,1991。263(3): 597 - 600页。[PubMed]
251.亨德里克森,。两栖动物视网膜的Landolt俱乐部:高尔基和电子显微镜研究。角膜切削投资,1966年。5(5): 484 - 96页。[IOVS]
252.脑,厦门市。非洲肺鱼视网膜上的Landolt 's club,原埃塞俄比亚肺鱼,Heckel。视觉Res, 1970。10(4): 299 - 306页。[PubMed]
253.Vigh, B.,等。视网膜的脑脊液接触神经元、感觉松果体细胞和Landolt 's俱乐部,通过对视蛋白的电子显微镜免疫反应显示。细胞组织研究,1983。233(3): 539 - 48页。[PubMed]
254.奎萨达和J.M.吉尼斯-加尔维斯,鸡视网膜Landolt 's club的形态和结构研究。J Morphol, 1985。184(2): 205 - 14页。[PubMed]
255.维,b,和i。维-泰希曼,接触csf的神经元和松果体细胞的细胞化学。《大脑研究》,1992年。91: 299 - 306页。[PubMed]
256.大卫,C,等人,在大西洋盲鳗的脑室系统中,脑脊液接触神经元。洪生物学报,2003。54(1): 35-44页。[PubMed]
257.García-Fernández, J.M.和R.G.福斯特,七鳃鳗幼虫下丘脑脑脊液接触神经元中感光蛋白的免疫细胞化学鉴定。细胞组织研究,1994。275(2): 319 - 326页。
258.科尔布,h·和e·v·费明列提,猫视网膜内丛状层的杆状和锥状通路。科学,1974。186: 47-9页。[PubMed]
259.纳尔逊,R。所有的无分泌细胞都能加快猫视网膜中杆状信号的传递时间。J Neurophysiol, 1982。47(5): 928 - 47页。[PubMed]
260.科尔布,h·r·纳尔逊,猫视网膜的神经结构。Retin Eye Res, 1984。3.: 21-60页。
261.斯特拉托伊、R.F.达歇和E.拉维奥拉,兔视网膜内丛状层杆状双极细胞的突触连接。神经科学杂志,1990。295(3): 449 - 66页。[PubMed]
262.范尼,d。i。h。m。杨,I.C.金瑟,兔视网膜中的杆状电路。Vis >, 1991。7(1 - 2): 141 - 54页。[PubMed]
263.斯特拉托伊,E·拉维奥拉,和r·f·达歇,兔视网膜内窄场双层棒无分泌细胞(AII)的突触连接。神经科学杂志,1992。325(2): 152 - 68页。[PubMed]
264.安石,J.F.和g。福尔克,角鲨黑暗适应视网膜中竿双极细胞的反应。杂志,1980年。300: 115 - 50页。[PubMed]
265.尼尔森,D.E.和S. Pelger,这是对眼睛进化所需时间的悲观估计。罗伊教授,1994。256: 53-8页。[PubMed]
266.D 'Aniello, S.等人,脊椎动物同源体基因Rx的海鞘同源体对眼细胞的发育和功能至关重要。分化,2006年。74(5): 222 - 34页。[PubMed]
267.蔡司,c.j.,等人,鸭嘴兽视网膜形态的比较。J Morphol, 2011。272(8): 949 - 57页。[PubMed]
268.伯恩斯,法医和T.D.兰姆,视杆和视锥感光器的视觉转导,在视觉神经科学, J.S. Werner和L.M. Chalupa,编辑。2004,麻省理工学院出版社:剑桥,马萨诸塞州。p . 215 - 233。
269.拉维奥拉,E.和N.B.吉鲁拉,视网膜外丛状层中特殊接触的膜内组织。猴子和兔子的冻裂研究。细胞生物学杂志,1975。65(1): 192 - 222页。[PubMed]
270.Rieke F.和D.A. Baylor,视网膜视锥细胞暗噪声的起源及其对功能的影响。神经元,2000年。26(1): 181 - 6页。[PubMed]
271.布热津斯基,j·和t·雷,视网膜组织发生,在眼睛百科全书。爱思唯尔有限公司.2010.p . 73 - 80。
272.戈麦斯,F.L.和M. Cayouette,视网膜发育:细胞类型规范,在神经科学百科全书L.R. Squire,编辑。2009,爱思唯尔/学术出版社:阿姆斯特丹;波士顿。p . 203 - 209。
273.陈志强,李旭东,C. Desplan,视网膜神经元规格的确定性或随机性选择。神经元,2012年。75(5): 739 - 42页。[PubMed]
274.瑞茜,B.E。,感光体的发育可塑性。大脑研究,2004年。144: p -。[PubMed]
275.兰姆,t.d.,小E.N.普,还有S.P.科林,脊椎动物眼睛的起源。进化:教育与推广,2008年。1(4): 415 - 426页。
276.Wassle、H。哺乳动物视网膜的平行处理。神经科学,2004。5(10): 747 - 57页。[PubMed]

作者
特雷弗·d·兰姆1971年在墨尔本大学获得电子工程学位,之后前往英国剑桥大学,在艾伦·霍奇金爵士的指导下进行研究生学习,与丹尼斯·贝勒一起研究视网膜水平细胞和光感受器。1975年获得博士学位。后来在美国斯坦福大学做博士后时,Denis Baylor, King-Wai Yau和Trevor开发了从光感受器记录的吸液移液技术。在接下来的25年里,他再次在英国剑桥大学从事光转导和光和暗适应方面的工作。1993年,他被选为皇家学会会员。与美国的Ed Pugh一起,他开发了一种数学描述,描述了光感受器光反应下的分子反应,以及黑暗适应和类视黄酮循环的模型。2003年,特雷弗回到澳大利亚,在堪培拉的澳大利亚国立大学(ANU)担任联邦研究员。自2011年以来,他一直是澳大利亚国立大学的荣誉教授,在那里他从事目前的兴趣在光感受器和视网膜的进化起源。电子邮件特在Trevor.Lamb@anu.edu.au